Распределение организмов по средам жизни
В процессе длительного исторического развития живой материи и формирования все более совершенных форм живых существ, организмы, осваивая новые места обитания, распределились на Земле соответственно ее минеральным оболочкам (гидросфера, литосфера, атмосфера) и приспособились к существованию в строго определенных условиях.
Первой средой жизни стала вода. Именно в ней возникла жизнь. По мере исторического развития многие организмы начали заселять наземно-воздушную среду. В результате появились наземные растения и животные, которые бурно эволюционировали, адаптируясь к новым условиям существования.
В процессе функционирования живого вещества на суше поверхностные слои литосферы постепенно преобразовывались в почву, в своеобразное, по выражению В. И. Вернадского, биокосное тело планеты. Почву стали заселять как водные, так и наземные организмы, создавая специфичный комплекс ее обитателей.
Таким образом, на современной Земле четко выделяются четыре среды жизни – водная, наземно-воздушная, почва и живые организмы,– существенно различающиеся своими условиями. Рассмотрим каждую из них.
Общая характеристика. Водная среда жизни, гидросфера, занимает до 71 % площади земного шара. По объему запасы воды на Земле исчисляются в пределах 1370 млн. куб. км, что составляет 1/800 часть объема земного шара. Основное количество воды, более 98 %, сосредоточено в морях и океанах, 1,24 % представлено льдами полярных областей; в пресных водах рек, озер и болот количество воды не превышает 0,45 %.
В водной среде обитает около 150000 видов животных (примерно 7 % от общего их количества на земном шаре) и 10 000 видов растений (8 %). Несмотря на то что представители абсолютного большинства групп растений и животных остались в водной среде (в своей «колыбели»), число их видов значительно меньше, чем наземных. Значит, эволюция на суше проходила значительно быстрее.
Наиболее разнообразен и богат растительный и животный мир морей и океанов экваториальных и тропических областей (особенно Тихого и Атлантического океанов). На юг и север от этих поясов качественный состав организмов постепенно обедняется. В районе Ост-Индского архипелага распространено около 40000 видов животных, а в море Лаптевых всего 400. При этом основная масса организмов Мирового океана сосредоточена в сравнительно небольшой по площади зоне морских побережий умеренного пояса и среди мангровых зарослей тропических стран. На обширных акваториях вдали от берега расположены пустынные области, практически лишенные жизни.
Удельный вес рек, озер и болот по сравнению с таковым морей и океанов в биосфере незначительный. Тем не менее они создают необходимый для огромного количества растений и животных, а также для человека запас пресной воды.
Водная среда оказывает сильное влияние на ее обитателей. В свою очередь живое вещество гидросферы воздействует на среду обитания, перерабатывает ее, вовлекая в круговорот веществ. Подсчитано, что вода морей и океанов, рек и озер разлагается и восстанавливается в биотическом круговороте за 2 млн. лет, т. е. вся она прошла через живое вещество планеты не одну тысячу раз *. Таким образом, современная гидросфера представляет собой продукт жизнедеятельности живого вещества не только современной, но и прошлых геологических эпох.
Характерной чертой водной среды является ее подвижность даже в стоячих водоемах, не говоря уже о проточных, быстро текущих реках и ручьях. В морях и океанах наблюдаются приливы и отливы, мощные течения, штормы; в озерах вода перемещается под действием ветра и температуры. Движение воды обеспечивает снабжение водных организмов кислородом и питательными веществами, приводит к выравниванию (снижению) температуры во всем водоеме.
Обитатели водоемов выработали соответствующие приспособления к подвижности среды. К примеру, в проточных водоемах встречаются прочно прикрепленные к подводным предметам так называемые «обрастающие» растения ·– зеленые водоросли (Cladophora) со шлейфом отростков, диатомовые (Diatomeae), водяные мхи (Fontinalis), образующие плотный покров даже на камнях в бурных речных перекатах.
Животные также адаптировались к подвижности водной среды. У рыб, обитающих в быстро текущих реках, тело в поперечном разрезе почти округлое (форель, гольян). Двигаются они обычно навстречу течению. Беспозвоночные проточных водоемов обычно держатся на дне, тело их сплющено в дорзо-вентральном направлении, многие имеют на брюшной стороне различные органы фиксации, позволяющие им прикрепляться к подводным предметам. В морях наиболее сильное влияние подвижных масс воды испытывают организмы приливно-отливной и прибойной зон. На скалистых берегах в прибойной полосе распространены прочно удерживающиеся на камнях усоногие раки (Balanus, Chthamalus), брюхоногие моллюски (Patella Haliotis), некоторые виды ракообразных, укрывающихся в расщелинах берега.
В жизни водных организмов умеренных широт большую роль играет вертикальное перемещение воды в стоячих водоемах. Вода в них четко делится на три слоя: верхний–эпилимнион, температура которого испытывает резкие сезонные колебания; слой температурного скачка– металимнион (термоклин), где наблюдается резкий перепад температур; придонный глубинный слой, гиполимнион,– здесь температура в течение года изменяется незначительно.
В летнее время наиболее теплые слои воды располагаются у поверхности, а холодные – у дна. Такое послойное распределение температур в водоеме называется прямой стратификацией. Зимой, с понижением температуры, наблюдается обратная стратификация: поверхностные холодные воды с температурой ниже 4 °С располагаются над сравнительно теплыми. Это явление называется температурной дихотомией. Оно особенно выражено в большинстве наших озер летом и зимой. В результате температурной дихотомии в водоеме образуется плотностная стратификация воды, нарушается вертикальная циркуляция ее и наступает период временного застоя –-стагнация.
Весной поверхностная вода вследствие нагревания до 4 °С становится более плотной и погружается вглубь, а на ее место с глубины поднимается более теплая вода. В результате такой вертикальной циркуляции в водоеме наступает гомотермия, т. е. на какое-то время температура всей водной массы выравнивается. С дальнейшим повышением температуры верхние слои воды становятся все менее плотными и уже не опускаются – наступает летняя стагнация.
Осенью же поверхностный слой охлаждается, становится более плотным и опускается вглубь, вытесняя на поверхность более теплую воду. Это происходит до наступления осенней гомотермии. При охлаждении поверхностных вод ниже 4 °С они снова становятся менее плотными и опять остаются на поверхности. В результате прекращается циркуляция воды и наступает зимняя стагнация.
Организмы в водоемах умеренных широт хорошо приспособлены к сезонным вертикальным перемещениям слоев воды, к весенней и осенней гомотермии и к летней и зимней стагнации (рис. 13).
В озерах тропических широт температура воды на поверхности никогда не опускается ниже 4 °С и температурный градиент в них четко выражен до самых глубинных слоев. Перемешивание воды, как правило, происходит здесь нерегулярно в наиболее холодное время года.
Своеобразные условия для жизни складываются не только в толще воды, но и на дне водоема, так как в грунтах отсутствует аэрация и из них вымываются минеральные соединения. Поэтому они не обладают плодородием и служат для водных организмов лишь более или менее твердым субстратом, выполняя в основном механико-динамическую функцию. В связи с этим наибольшее экологическое значение приобретают размеры частиц грунта, плотность их прилегания друг к другу и устойчивость к смыву течениями.
Абиотические факторы водной среды. Вода как среда жизни обладает особыми физико-химическими свойствами.
Температурный режим гидросферы коренным образом отличается от такового в других средах. Колебания температур в Мировом океане сравнительно невелики: самая низкая около –2 °С, а самая высокая приблизительно 36 °С. Амплитуда колебаний здесь, таким образом, укладывается в 38 °С. С глубиной температура воды в океанах падает. Даже в тропических областях на глубине 1000 м не превышает 4–5°С. На глубинах всех океанов залегает слой холодной воды (от -1,87 до +2°С).
В пресных внутренних водоемах умеренных широт температура поверхностных слоев воды колеблется от – 0,9 до +25°С, в более глубинных составляет 4–5 °С. Исключением являются термальные источники, где температура поверхностного слоя достигает иногда 85– 93 °С.
Такие термодинамические особенности водной среды, как высокая удельная теплоемкость, большая теплопроводность и расширение при замерзании, создают особо благоприятные условия для жизни. Эти условия обеспечиваются и высокой скрытой теплотой плавления воды, в результате чего зимой подо льдом температура никогда не бывает ниже точки замерзания ее (для пресных вод около 0°С). Так как наибольшей плотностью вода обладает при 4° С, а при замерзании расширяется, то зимой лед образуется лишь сверху, основная же толща не промерзает.
Поскольку температурный режим водоемов характеризуется большой стабильностью, организмы, обитающие в ней, отличаются относительным постоянством температуры тела и обладают узким диапазоном приспособленности к колебаниям температуры среды. Даже незначительные отклонения в тепловом режиме се могут привести к существенным изменениям в жизни животных и растений. Примером может служить «биологический взрыв» лотоса (Nelumbium caspium) в самой северной части его обитания – в дельте Волги. Длительное время это экзотическое растение заселяло лишь небольшой залив. За последнее десятилетие площадь лотосовых зарослей увеличилась почти в 20 раз и занимает теперь свыше 1500 га акватории. Такое бурное распространение лотоса объясняется общим падением уровня Каспийского моря, что сопровождалось образованием множества мелких озер и лиманов в устье Волги. В жаркие летние месяцы вода здесь прогревалась сильнее, чем прежде, это и способствовало разрастанию зарослей лотоса.
Воде свойственна также значительная плотность (в этом отношении она в 800 раз превосходит воздушную среду) и вязкость. На растениях эти особенности сказываются в том, что у них очень слабо или вовсе не развивается механическая ткань, поэтому стебли их очень эластичны и легко изгибаются. Большинству водных растений присуща плавучесть и способность находиться в толще воды во взвешенном состоянии. Они то поднимаются к поверхности, то вновь опускаются. У многих водных животных покровы обильно смазываются слизью, уменьшающей трение при передвижении, а тело приобретает обтекаемую форму.
Организмы в водной среде распределены по всей ее толще (в океанических впадинах животные обнаружены на глубинах свыше 10000 м). Естественно, что на разных глубинах они испытывают различное давление. Глубоководные приспособлены к высокому давлению (до 1000 атм), обитатели же поверхностных слоев ему не подвержены. В среднем в водной толще на каждые 10 м глубины давление возрастает на 1 атм. Все гидробионты приспособлены к этому фактору и соответственно делятся на глубоководных и обитающих на незначительных глубинах.
Большое влияние на водные организмы оказывают прозрачность воды и ее световой режим. Особенно это сказывается на распространении фотосинтезирующих растений. В мутных водоемах они обитают только в поверхностном слое, а там, где большая прозрачность, проникают на значительные глубины. Определенную мутность воды создает огромное количество взвешенных в ней частиц, что лимитирует проникновение солнечных лучей. Мутность воды может быть обусловлена частицами минеральных веществ (глина, ил), мелкими организмами. Уменьшается прозрачность воды и летом при бурном разрастании водной растительности, при массовом размножении мелких организмов, находящихся во взвешенном состоянии в поверхностных слоях. Световой режим водоемов зависит и от сезона. На севере в умеренных широтах, когда водоемы замерзают, а лед сверху еще покрывается снегом, проникновение света в толщу воды сильно ограничивается.
Световой режим обусловливается также закономерным убыванием света с глубиной из-за того, что вода поглощает солнечный свет. При этом лучи с разной длиной волны поглощаются неодинаково: быстрее всего–красные, тогда как сине-зеленые проникают на значительные глубины. В океане с глубиной становится все темнее. Цвет среды при этом меняется, постепенно переходя от зеленоватого до зеленого, затем до голубого, синего, сине-фиолетового, сменяемого постоянным мраком. Соответственно этому с глубиной зеленые водоросли (Chlorophyta) сменяются бурыми (Phaeophyta) и красными (Rhodophyta), пигменты которых приспособлены к улавливанию солнечных лучей с разной длиной волны. С глубиной также закономерно меняется и окраска животных. В поверхностных, светлых слоях воды обычно обитают ярко и разнообразно окрашенные животные, тогда как глубоководные виды лишены пигментов. В сумеречной зоне океана обитают животные, окрашенные в цвета с красноватым оттенком, что помогает им скрываться от врагов, так как красный цвет в сине-фиолетовых лучах воспринимается как черный.
Немаловажную роль в жизни водных организмов играет соленость воды. Как известно, вода является прекрасным растворителем многих минеральных соединений. В результате природным водоемам свойствен определенный химический состав. Наибольшее значение имеют карбонаты, сульфаты, хлориды. Количество растворенных солей на 1 л воды в пресных водоемах не превышает 0,5 г (обычно меньше), в морях и океанах оно достигает 35 г (табл. 6).
Таблица 6. Распределение основных солей в различных водоемах (по Р. Дажо, 1975)
Для жизни пресноводных животных существенную роль играет кальций. Моллюски, ракообразные и другие беспозвоночные используют его для построения раковин, наружного скелета. Но пресные водоемы в зависимости от целого ряда обстоятельств (наличие тех или иных растворимых солей в грунте водоема, в грунте и почве берегов, в воде впадающих рек и ручьев) очень различаются как по составу, так и по концентрации растворенных в них солей. Морские воды более стабильны в этом отношении. В них обнаружены почти все известные элементы. Однако по степени значимости первое место занимает поваренная соль, затем хлористый и сернокислый магний и хлористый калий.
Пресноводные растения и животные обитают в гипотонической среде, т. е. в среде, в которой концентрация растворенных веществ ниже, чем в жидкостях тела и тканей. Из-за разницы в осмотическом давлении вне и внутри тела в организм постоянно проникает вода, и гидробионты пресных вод вынуждены интенсивно удалять ее. В связи с этим у них хорошо выражены процессы осморегуляции. Концентрация солей в жидкостях тела и тканей многих морских организмов изотонична концентрации растворенных солей в окружающей воде. Поэтому осморегуляторные функции у них не развиты в такой степени, как у пресноводных. Трудности осморегуляции являются одной из причин того, что многие морские растения и особенно животные не сумели заселить пресные водоемы и оказались, за исключением отдельных представителей, типичными морскими жителями (кишечнополостные – Coelenterata, иглокожие–·Echinodermata, погонофоры – Pogonophora, губки – Spongia, оболочники – Tunicata). В то же время в морях и океанах практически не обитают насекомые, тогда как пресноводные бассейны обильно ими заселены. Типично морские и типично пресноводные виды не переносят значительных изменений солености воды. Все они – стеногалинные организмы. Эвригалинных животных пресноводного и морского происхождения сравнительно немного. Обычно они встречаются, притом в значительных количествах, в солоноватых водах. Это пресноводный судак (Stizostedion lucioperca), лещ (Abramis brama), щука (Esox lucius), из морских можно назвать семейство кефалевых (Mugilidae).
В пресных водах распространены растения, укрепленные на дне водоема. Нередко их фотосинтетическая поверхность располагается над водой. Это рогозы (Typha), камыши (Scirpus), стрелолист (Sagittaria), кувшинки (Nymphaea), кубышки (Nuphar). У других фотосинтезирующие органы погружены в воду. К ним относятся рдесты (Potamogeton), уруть (Myriophyllum), элодея (Elodea). Некоторые высшие растения пресных вод лишены корней. Они либо свободно плавают, либо обрастают подводные предметы или водоросли, прикрепленные к грунту.
Если кислород для воздушной среды не играет существенной роли, то для водной – это важнейший экологический фактор. Содержание его в воде обратно пропорционально температуре. С понижением температуры растворимость кислорода, как и других газов, увеличивается. Накопление растворенного в воде кислорода происходит в результате поступления его из атмосферы, а также вследствие фотосинтетической деятельности зеленых растений. При перемешивании воды, что характерно для проточных водоемов и особенно для быстро текущих рек и ручьев, содержание кислорода также возрастает.
Различные животные проявляют неодинаковую потребность к кислороду. Например, форель (Salmo trutta), гольян (Phoxinus phoxinus) очень чувствительны к его дефициту и поэтому обитают лишь в быстро текущих холодных и хорошо перемешиваемых водах. Плотва (Rutilus rutilus), ерш (Acerina cernua), сазан (Cyprinus carpio), карась (Carassius carassius) неприхотливы в этом отношении, а личинки комаров хирономид (Chirono-midae) и малощетинковые черви трубочники (Tubifex) обитают на больших глубинах, где кислород вообще отсутствует или его очень мало. Водные насекомые и легочные моллюски (Pulmonata) также могут жить в водоемах с низким содержанием кислорода. Однако они систематически поднимаются к поверхности, запасая на некоторое время свежий воздух.
Углекислый газ растворяется в воде примерно в 35 раз лучше кислорода. В воде его почти в 700 раз больше, чем в атмосфере, откуда он поступает. Источником углекислоты в воде, кроме того, являются карбонаты и бикарбонаты щелочных и щелочноземельных металлов. Углекислый газ, содержащийся в воде, обеспечивает фотосинтез водных растений и принимает участие в формировании известковых скелетных образований беспозвоночных животных.
Большое значение в жизни водных организмов имеет концентрация водородных ионов (рН). Пресноводные бассейны с рН 3,7–4,7 считаются кислыми, 6,95–7,3 – нейтральными, с рН больше 7,8 – щелочными. В пресных водоемах рН испытывает даже суточные колебания. Морская вода более щелочная и рН ее значительно меньше изменяется, чем пресной. С глубиной рН уменьшается.
Концентрация водородных ионов играет большую роль в распределении гидробионтов. При рН меньше 7,5 растет полушник (Isoetes), ежеголовник (Sparganium), при 7,7–8,8, т. е. в щелочной среде, развиваются многие виды рдестов, элодея. В кислых водах болот преобладают сфагновые мхи (Sphagnum), но отсутствуют пластинчатожаберные моллюски из рода беззубок (Unio), редко встречаются другие моллюски, но в обилии распространены раковинные корненожки (Testacea). Большинство пресноводных рыб выдерживают рН от 5 до 9. Если рН меньше 5, наблюдается массовая гибель рыб, а выше 10 – погибают все рыбы и другие животные.
Экологические группы гидробионтов. Толща воды - пелагиаль (pelagos – море) заселена пелагическими организмами, способными активно плавать или удерживаться (парить) в определенных слоях. В соответствии с этим пелагические организмы подразделяются на две группы – нектон и планктон. Обитатели дна образуют третью экологическую группу организмов – бентос.
Нектон (nekios –· плавающий) – это совокупность пелагических активно передвигающихся животных, не имеющих непосредственной связи с дном. В основном это крупные животные, способные преодолевать большие расстояния и сильные водные течение. Для них характерна обтекаемая форма тела и хорошо развитые органы движения. Типичными нектонными организмами являются рыбы, кальмары, ластоногие, киты. В пресных водах, кроме рыб, к нектону относятся земноводные и активно перемещающиеся насекомые. Многие морские рыбы могут передвигаться в толще воды с огромной скоростью. Очень быстро, до 45–50 км/ч, плавают некоторые кальмары (Oegopsida), парусники (Istiopharidae) развивают скорость до 100–ПО км/ч, а меч-рыба (Xiphias glabius) – до 130 км/ч.
Планктон (planktos – парящий, блуждающий) – это совокупность пелагических организмов, не обладающих способностью к быстрым активным передвижениям. Планктонные организмы не могут противостоять течениям. В основном это мелкие животные – зоопланктон и растения – фитопланктон. В состав планктона периодически включаются и парящие в толще воды личинки многих животных.
Планктонные организмы располагаются либо на поверхности воды, либо на глубине или даже в придонном слое. Первые составляют особую группу – нейстон. Организмы же, часть тела которых находится в воде, а часть – над ее поверхностью, получили название плейстон. Таковы сифонофоры (Siphonophora), ряска (Lemna) и др.
Фитопланктон имеет большое значение в жизни водоемов, поскольку является основным продуцентом органического вещества. К нему относятся прежде всего диатомовые (Diatomeae) и зеленые (Chlorophyta) водоросли, растительные жгутиконосцы (Phytomastigina), перидинеи (Peridineae) и кокколитофориды (Coccolitophoridae). В северных водах Мирового океана преобладают диатомовые водоросли, а в тропических и субтропических – панцирные жгутиконосцы. В пресных водах, кроме диатомовых, распространены зеленые и сине-зеленые (Суаnophyta) водоросли.
Зоопланктон и бактерии встречаются на всех глубинах. В морском зоопланктоне доминируют мелкие ракообразные (Copepoda, Amphipoda, Euphausiacea), простейшие (Foraminifera, Radiolaria, Tintinnoidea). Более крупные представители его – это крылоногие моллюски (Pteropoda), медузы (Scyphozoa) и плавающие гребневики (Ctenophora), сальпы (Salpae), некоторые черви (Alciopidae, Tomopteridae). В пресных водах распространены плохо плавающие сравнительно крупные ракообразные (Daphnia, Cyclopoidea, Ostracoda, Simocephalus; рис. 14), много коловраток (Rotatoria) и простейших.
Наибольшего видового разнообразия достигает планктон тропических вод.
Группы планктонных организмов различают по размеру. Наннопланктон (nannos – карликовый) наиболее мелкие водоросли и бактерии; микропланктон (micros – малый) – большинство водорослей, простейшие, коловратки; мезопланктон (mеsos – средний) – веслоногие и ветвистоусые раки, креветки и ряд животных и растений, не больше 1 см в длину; макропланктон (macros – крупный) –·медузы, мизиды, креветки и другие организмы, размером более 1 см; мегалопланктон (megalos– громадный) – очень крупные, свыше 1 м, животные. Например, плавающий гребневик венерин пояс (Cestus veneris) достигает в длину 1,5 м, а медуза цианея (Суапеа) имеет колокол диаметром до 2 м и щупальца длиною в 30 м.

Планктонные организмы являются важным пищевым компонентом многих водных животных (в том числе таких гигантов, как усатые киты – Mystacoceti), особенно если учесть, что для них, и прежде всего для фитопланктона, характерны сезонные вспышки массового размножения (цветение воды).
Бентос (benthos – глубина) – совокупность организмов, обитающих на дне (на грунте и в грунте) водоемов. Он подразделяется на фитобентос и зообентос. В основном представлен прикрепленными или медленно передвигающимися, а также роющимися в грунте животными. Только на мелководье он состоит из организмов, синтезирующих органическое вещество (продуценты), потребляющих (консументы) и разрушающих (редуценты) его. На больших глубинах, куда не проникает свет, фитобентос (продуценты) отсутствует.
Бентосные организмы различаются по образу жизни – подвижные, малоподвижные и неподвижные; по способу питания – фотосинтезирующие, плотоядные, растительноядные, детритоядные; по размерам – макро-, мезо- микробентос.
Фитобентос морей в основном включает бактерии и водоросли (диатомовые, зеленые, бурые, красные). У побережий встречаются также цветковые растения: зостера (Zostera), филлосподикс (Phyllospadix), руппия (Rup-pia). Наиболее богат фитобентос на скалистых и каменистых участках дна. У побережий ламинарии (Laminaria) и фукусы (Fucus) иногда образуют биомассу до 30 кг на 1 кв. м. На мягких грунтах, где растения не могут прочно прикрепляться, фитобентос развивается в основном в защищенных от волн местах.
Фитобенос пресных вод представлен бактериями, диатомовыми и зелеными водорослями. Обильны прибрежные растения, располагающиеся от берега вглубь четко выраженными поясами. В первом поясе произрастают полупогруженные растения (тростники, камыши, рогоз и осоки). Второй пояс занимают погруженные растения с плавающими листьями (кубышки, кувшинки, ряски, водокрас). В третьем поясе преобладают погруженные растения – рдесты, элодея и др.
Все водные растения по образу жизни можно разделить на две основные экологические группы: гидрофиты – растения, погруженные в воду только нижней частью и обычно укореняющиеся в грунте, и гидатофиты – растения, полностью погруженные в воду, но иногда плавающие на поверхности или имеющие плавающие листья.
В морском зообентосе доминируют фораминиферы, губки, кишечнополостные, немертины, многощетинковые черви, сипункулиды, мшанки, плеченогие, моллюски, асцидии, рыбы. Наиболее многочисленны бентосные формы на мелководьях, где общая биомасса их нередко достигает десятков килограммов на 1 кв. м. С глубиной численность бентоса резко падает и на больших глубинах составляет миллиграммы на 1 кв. м.
В. пресных водоемах зообентоса меньше, чем в морях и океанах, а видовой состав однообразнее. В основном это простейшие, некоторые губки, ресничные и малощетинковые черви, пиявки, мшанки, моллюски и личинки насекомых.
Экологическая пластичность водных организмов. Водные организмы обладают меньшей экологической пластичностью, чем наземные, так как вода – более стабильная среда и абиотические факторы ее претерпевают сравнительно незначительные колебания. Наименее пластичны морские растения и животные. Они очень чувствительны к изменениям солености воды и ее температуры. Так, мадрепоровые кораллы не выдерживают даже слабого опреснения воды и обитают только в морях, притом на твердом грунте при температуре не ниже 20 °С. Это типичные стенобионты. Однако есть виды и с повышенной экологической пластичностью. Например, корненожка Cyphoderia ampulla является типичным эврибионтом. Она обитает в морях и в пресных водах, в теплых прудах и в холодных озерах.
Пресноводные животные и растения, как правило, гораздо пластичнее, чем морские, поскольку пресная вода как среда жизни более изменчива. Самыми же пластичными являются солоноватоводные обитатели. Они адаптированы и к высокой концентрации растворенных солей и к значительному опреснению. Однако их насчитывается сравнительно небольшое количество видов, поскольку в солоноватых водах экологические факторы претерпевают существенные изменения.
Широту экологической пластичности гидробионтов оценивают по отношению не только ко всему комплексу факторов (эври- и станобионтность), но и к какому-либо одному из них. Прибрежные растения и животные в отличие от обитателей открытых зон в основном эвритермные и эвригалинные организмы, так как вблизи берега температурные условия и солевой режим довольно изменчивы (прогреваемость солнцем и сравнительно интенсивное охлаждение, опреснение притоком воды из ручьев и рек, особенно в период дождей, и т. д.). Типичным стенотермным видом является лотос. Он произрастает только в хорошо прогреваемых мелких водоемах. По этим же причинам более эвритермными и эвригалинными оказываются обитатели поверхностных слоев в сравнении с глубоководными формами.
Экологическая пластичность служит важным регулятором расселения организмов. Как правило, гидробионты с высокой экологической пластичностью распространены довольно широко. Это касается, к примеру, элодеи. Однако диаметрально противоположен ей в этом смысле рачок артемия (Artemia salina). Он живет в небольших водоемах с очень соленой водой. Это типичный стеногалинный представитель с узкой экологической пластичностью. Но по отношению к другим факторам он весьма пластичен и поэтому в соленых водоемах встречается повсеместно.
Экологическая пластичность зависит от возраста и фазы развития организма. Так, морской брюхоногий моллюск Littorina во взрослом состоянии ежедневно при отливах длительное время обходится без воды, а его личинки ведут сугубо планктонный образ жизни и не переносят высыхания.
Адаптивные особенности водных растений. Экология водных растений, как было отмечено, весьма специфична и резко отличается от экологии большинства наземных растительных организмов. Способность водных растений поглощать влагу и минеральные соли непосредственно из окружающей среды отражается на их морфологической и физиологической организации. Для водных растений прежде всего характерно слабое развитие проводящей ткани и корневой системы. Последняя служит в основном для прикрепления к подводному субстрату и в отличие от наземных растений не выполняет функцию минерального питания и водоснабжения. В связи с этим корни укореняющихся водных растений лишены корневых волосков. Питание их осуществляется всей поверхностью тела. Мощно развитые корневища у некоторых из них служат для вегетативного размножения и запасания питательных веществ. Таковы многие рдесты, кувшинки, кубышки.
Большая плотность воды обусловливает возможность обитания растений во всей ее толще. Для этого у низших растений, заселяющих различные слои и ведущих плавающий образ жизни, имеются специальные придатки, увеличивающие их плавучесть и позволяющие им удерживаться во взвешенном состоянии. У высших гидрофитов слабо развивается механическая ткань. В их листьях, стеблях, корнях, как отмечалось, располагаются воздухоносные межклеточные полости. Это увеличивает легкость и плавучесть взвешенных в воде и плавающих на поверхности органов, а также способствует смыванию внутренних клеток водой с растворенными в ней газами и солями. Гидатофиты в целом характеризуются большой поверхностью листьев при незначительном общем объеме растения. Это обеспечивает им интенсивный газообмен при недостатке растворенного в воде кислорода и других газов. У многих рдестов (Potamogeton lusens, P. perfoliatus) стебли и листья тонкие и очень длинные, покровы их легко проницаемы для кислорода. Другие растения обладают сильно расчлененными листьями (водяной лютик– Ranunculus aquatilis, уруть – Myriophyllum spicatum, роголистник – Ceratophyllum dernersum).
У ряда водных растений развита гетерофилия (разнолистность). Например, у сальвинии (Salvinia) погруженные листья выполняют функцию минерального питания, а плавающие – органического. У кувшинки и кубышки плавающие и погруженные в воду листья существенно отличаются друг от друга. Верхняя поверхность плавающих листьев плотная и кожистая с большим количеством устьиц. Это способствует лучшему газообмену с воздухом. На нижней стороне плавающих и на подводных листьях устьиц совсем нет.
Не менее важной адаптивной особенностью растений к обитанию в водной среде является и то, что погруженные в воду листья обычно очень тонкие. Хлорофилл в них часто располагается в клетках эпидермиса. Это приводит к усилению интенсивности фотосинтеза в условиях слабого освещения. Наиболее четко такие анатомо-морфологические особенности выражены у многих рдестов (Potamogeton), элодеи (Helodea canadensis), водяных мхов (Riccia, Fontinalis), валиснерии (Vallisneria spiralis).
Защитой водных растений от вымывания из клеток минеральных солей (выщелачивания) является выделение специальными клетками слизи и образование эндодермы в виде кольца из более толстостенных клеток.
Сравнительно низкая температура водной среды обусловливает отмирание вегетирующих частей у погруженных в воду растений после образования зимних почек, а также замену летних нежных тонких листьев более жесткими и короткими зимними. Вместе с тем низкая температура воды отрицательно влияет на генеративные органы водных растений, а высокая ее плотность затрудняет перенос пыльцы. Поэтому водные растения интенсивно размножаются вегетативным путем. Половой процесс у многих из них подавлен. Адаптируясь к особенностям водной среды, большинство погруженных и плавающих на поверхности растений выносят цветоносные стебли в воздушную среду и размножаются половым путем (пыльца разносится ветром и поверхностными течениями). Образующиеся плоды, семена и другие зачатки также распространяются поверхностными течениями (гидрохория).
К гидрохорным принадлежат не только водные, но и многие прибрежные растения. Плоды их обладают высокой плавучестью и могут длительное время находиться в воде, не теряя всхожести. Водой переносятся плоды и семена частухи (Alisma plantago-aquatica), стрелолиста (Sagittaria sagittifolia),сусака (Butomusumbellatus), рдестов и других растений. Плоды многих осок (Сагех) заключены в своеобразные мешочки с воздухом и тоже разносятся водными течениями. Предполагают, что даже кокосовые пальмы расселились по архипелагам тропических островов Тихого океана благодаря плавучести своих плодов – кокосовых орехов. Вдоль реки Вахш по каналам таким же образом расселился сорняк гумай (Sorgnum halepense).
Адаптивные особенности водных животных. Адаптации животных к водной среде еще более многообразны, чем растений. У них можно выделить анатомо-морфологические, физиологические, поведенческие и другие адаптивные признаки. Даже простое перечисление их затруднительно. Поэтому назовем в общих чертах лишь наиболее характерные из них.
Животные, обитающие в толще воды, обладают прежде всего приспособлениями, увеличивающими их плавучесть и позволяющими им противостоять движению воды, течениям. Донные организмы, наоборот, вырабатывают приспособления, препятствующие поднятию их в толщу воды, т. е. уменьшающие плавучесть и позволяющие удерживаться на дне даже в быстро текущих водах.
У мелких форм, живущих в толще воды, наблюдается редукция скелетных образований. У простейших (Rhizopoda, Radiolaria) раковины обладают пористостью, кремневые иглы скелета полые внутри. Удельная плотность медуз (Scyphozoa) и гребневиков (Ctenophora) уменьшается за счет наличия воды в тканях. Увеличение плавучести достигается и скоплением капелек жира в теле (ночесветки – Noctiluca, радиолярии – Radiolaria). Более крупные скопления жира наблюдаются и у некоторых ракообразных (Cladocera, Copepoda), рыб, китообразных. Удельную плотность тела снижают и газовые пузырьки в протоплазме раковинных амеб, воздухоносные камеры в раковинах моллюсков. Плавательные пузыри, наполненные газом, имеют многие рыбы. У сифонофор Physalia и Velella развиваются мощные воздухоносные полости.
Для пассивно плавающих в толще воды животных характерно не только уменьшение веса, но и увеличение удельной поверхности тела. Дело в том, что чем больше вязкость среды и выше удельная поверхность тела организма, тем медленнее он погружается в воду. В результате у животных уплощается тело, на нем образуются всевозможные шипы, выросты, придатки. Это свойственно многим радиоляриям (Chalengeridae, Aulacantha), жгутиковым (Leptodiscus, Craspedotella), фораминиферам (Globigerina, Orbulina). Поскольку с повышением температуры вязкость воды уменьшается, а с повышением солености – увеличивается, адаптации к увеличению трения наиболее резко выражены при высоких температурах и низких соленостях. Например, жгутиковые Ceratium из Индийского океана вооружены более длинными роговидными придатками, чем те, которые обитают в холодных водах Восточной Атлантики.
Активное плавание у животных осуществляется с помощью ресничек, жгутиков, изгибания тела. Так передвигаются простейшие, ресничные черви, коловратки.
Среди водных животных распространено плавание реактивным способом за счет энергии выбрасываемой струи воды. Это характерно для простейших, медуз, личинок стрекоз, некоторых двустворчатых моллюсков. Наивысшего совершенства реактивный способ передвижения достигает у головоногих моллюсков. Некоторые кальмары при выбрасывании воды развивают скорость 40–50 км/ч. У более крупных животных образуются специализированные конечности (плавательные ноги у насекомых, ракообразных; плавники, ласты). Тело у таких животных покрыто слизью и имеет обтекаемую форму.
Большая группа животных, в основном пресноводных, при передвижении использует поверхностную пленку воды (поверхностное натяжение). По ней свободно бегают, например, жуки вертячки (Gyrinidae), клопы водомерки (Gerridae, Veliidae). По нижней поверхности пленки передвигаются мелкие жуки Hydrophilidae, к ней подвешиваются и моллюски прудовики (Limnaea), личинки комаров. Все они обладают рядом особенностей в строении конечностей, а покровы их не смачиваются водой.
Только в водной среде встречаются неподвижные, ведущие прикрепленный образ жизни животные. Им свойственны своеобразная форма тела, незначительная плавучесть (плотность тела больше плотности воды) и специальные приспособления для прикрепления к субстрату. Одни прикрепляются к грунту, другие ползают по нему или ведут роющий образ жизни, некоторые поселяются на подводных предметах, в частности днищах кораблей.
Из прикрепленных к грунту животных наиболее характерны губки, многие кишечнополостные, особенно гидроиды (Hydroidea) и коралловые полипы (Anthozoa), морские лилии (Crinoidea), двустворчатые моллюски (Bivalvia), усоногие раки (Cirripedia) и др.
Среди роющих животных особенно много червей, личинок насекомых, а также моллюсков. Значительное время в грунте проводят определенные рыбы (шиповка – Cobitis taenia, камбаловые – Pleuronectidae, скаты – Rajidae), личинки миног (Petromyzones). Обилие этих животных и их видовое разнообразие зависят от типа грунта (камни, песок, глина, ил). На каменистых грунтах их обычно меньше, чем на илистых. Беспозвоночные, в массе заселяющие илистые грунты, создают оптимальные условия для жизни ряда более крупных придонных хищников.
Большинство водных животных пойкилотермны, и температура их тела зависит от температуры окружающей среды. У гомойотермных же млекопитающих (ластоногие, китообразные) образуется мощный слой подкожного жира, выполняющий теплоизоляционную функцию.
Для водных животных имеет значение давление среды обитания. В связи с этим выделяются стенобатные животные, не выдерживающие больших колебаний давления, и эврибатные, обитающие и при высоком и при низком давлении. Голотурии (Elpidia, Myriotrochus) живут на глубине от 100 до 9000 м, а многие виды раков Storthyngura, погонофоры, морские лилии располагаются на глубинах от 3000 до 10 000 м. У таких глубоководных животных отмечаются специфичные черты организации: увеличение размеров тела; исчезновение или слабое развитие известкового скелета; часто – редукция органов зрения; усиление развития осязательных рецепторов; отсутствие пигментации тела или, наоборот, темная окраска.
Поддержание определенного осмотического давления и ионного состояния растворов в теле животных обеспечивается сложными механизмами водно-солевого обмена. Однако большинство водных организмов пойкилосмотичны, т. е. осмотическое давление в их теле зависит от концентрации растворенных солей в окружающей воде. Гомойосмотичны лишь позвоночные, высшие раки, насекомые и их личинки – они сохраняют постоянное осмотическое давление в теле, независимо от солености воды.
Морские беспозвоночные в основном не имеют механизмов водно-солевого обмена: анатомически они закрыты для воды, а осмотически открыты. Однако говорить об абсолютном отсутствии у них механизмов, управляющих водно-солевым обменом, было бы неверно.

Они просто несовершенны, и это объясняется тем, что соленость морской воды близка к солености соков тела. Ведь у гидробионтов пресных вод соленость и ионное состояние минеральных веществ соков тела, как правило, выше, чем окружающей воды. Поэтому у них хорошо выражены механизмы осморегуляцни. Наиболее распространенным способом поддержания постоянного осмотического давления является регулярное удаление поступающей в организм воды с помощью пульсирующих вакуолей и органов выделения. У других животных для этих целей развиваются непроницаемые покровы из хитина или роговых образований. У некоторых на поверхности тела выделяется слизь.
Трудностью регулирования осмотического давления у пресноводных организмов объясняется их видовая бедность по сравнению с обитателями моря.
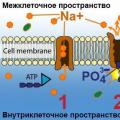
Проследим на примере рыб, как осуществляется осморегуляция животных в морских и пресных водах. Пресноводные рыбы избыток воды удаляют усиленной работой выделительной системы, а соли поглощают через жаберные лепестки. Морские рыбы, наоборот, вынуждены пополнять запасы воды и поэтому пьют морскую воду, а излишки поступающих с ней солей выводят из организма через жаберные лепестки (рис. 15).
Смена условий в водной среде вызывает определенные поведенческие реакции организмов. С изменением освещенности, температуры, солености, газового режима и других факторов связаны вертикальные миграции животных. В морях и океанах в таких миграциях (опускание вглубь, поднятие к поверхности) принимают участие миллионы тонн гидробионтов. При горизонтальных миграциях водные животные могут преодолевать сотни и тысячи километров. Таковы нерестовые, зимовальные и нагульные миграции многих рыб, водных млекопитающих.
Биофильтраторы и их экологическая роль. Одной из специфических особенностей водной среды является наличие в ней большого количества мелких частиц органического вещества – детрита, образующегося за счет отмирающих растений и животных. Огромные массы этих частиц оседают на бактериях и благодаря выделяющемуся в результате бактериального процесса газу постоянно находятся в толще воды во взвешенном состоянии.
Детрит для многих водных организмов представляет собой высококачественную пищу, поэтому некоторые из них, так называемые биофильтраторы, приспособились добывать его с помощью специфических микропористых структур. Эти структуры как бы отцеживают воду, задерживая взвешенные в ней частицы. Такой способ питания называется фильтрацией. Другая группа животных осаждает детрит на поверхность либо собственного тела, либо на специальные ловчие устройства. Такой способ называется седиментацией. Часто один и тот же организм питается путем и фильтрации и седиментации.
Животные-биофильтраторы (пластинчатожаберные моллюски, сидячие иглокожие и многощетинковые кольчецы, мшанки, асцидии, планктонные ракообразные и многие другие) играют большую роль в биологической очистке водоемов. Например, колония мидий (Mytilus) на 1 кв. м пропускает через мантийную полость до 250 куб. м воды в сутки, профильтровывая ее и осаждая взвешенные частицы. Почти микроскопический рачок каланус (Calanoida) очищает в сутки до 1,5 л воды. Если учесть громадную численность этих ракообразных, то работа, выполняемая ими по биологической очистке водоемов, представляется поистине грандиозной.
В пресных водах активными биофильтраторами являются перловицы (Unioninae), беззубки (Anodontinae), дрейссены (Dreissena), дафнии (Daphnia) и другие беспозвоночные. Значение их как своеобразной биологической «очистительной системы» водоемов настолько велико, что переоценить его практически невозможно.
Зональность водной среды. Для водной среды жизни характерна четко выраженная горизонтальная и особенно вертикальная зональность. Все гидробионты строго приурочены к обитанию в определенных зонах, различающихся различными условиями обитания.
В Мировом океане толща воды носит название пелагиаль, а дно – бенталь. Соответственно различают и экологические группы организмов, обитающих в толще воды (пелагические) и на дне (бентосные).
Дно в зависимости от глубины его залегания от поверхности воды, делится на сублитораль (область плавного понижения до глубины 200 м), батиаль (крутой склон), абиссаль (океаническое ложе со средней глубиной 3–6 км), ультраабиссаль (дно океанических впадин, расположенное на глубине от 6 до 10 км). Выделяется еще литораль – кромка берега, периодически заливаемая во время приливов (рис. 16).
Открытые воды Мирового океана (пелагиаль) также делятся на зоны по вертикали соответственно зонам бентали: эпипелагиаль, батипелагиаль, абиссопелагиаль.
Наиболее богато растениями и животными заселены литоральная и сублиторальная зоны. Здесь много солнечного света, низкое давление, значительные колебания температуры. Обитатели абиссальных и ультраабиссальных глубин живут при постоянной температуре, в темноте, испытывают громадное давление, достигающее в океанических впадинах нескольких сотен атмосфер.

Подобная, но менее четко выраженная зональность характерна и для внутренних пресных водоемов.
Водная среда включает поверхностные и подземные воды. Поверхностные воды в основном сосредоточены в океане, содержанием 1 млрд. 375 млн. км 3 - около 98% всей воды на Земле. Поверхность океана (акватория) составляет 361 млн. км 2 . Она примерно в 2,4 раза больше площади суши территории, занимающей 149 млн. км 2 . Вода в океане соленая, причем большая ее часть (более 1 млрд. км 3) сохраняет постоянную соленость около 3,5% и температуру, примерно равную 3,7°С. Заметные различия в солености и температуре наблюдаются почти исключительно в поверхностном слое воды, а также в окраинных и особенно в средиземных морях. Содержание растворенного кислорода в воде существенно уменьшается на глубине 50-60 метров.
Подземные воды бывают солеными, солоноватыми (меньшей солености) и пресными; существующие геотермальные воды имеют повышенную температуру (более 30°С.). Для производственной деятельности человечества и его хозяйственно-бытовых нужд требуется пресная вода, количество которой составляет всего лишь 2,7% общего объема воды на Земле, причем очень малая ее доля (всего 0,36%) имеется в легкодоступных для добычи местах. Большая часть пресной воды содержится в снегах и пресноводных айсбергах, находящихся в районах в основном Южного полярного круга. Годовой мировой речной сток пресной воды составляет 37,3 тыс. км 3 . томе того, может использоваться часть подземных вод, равная 13 тыс. км 3 . К сожалению, большая часть речного стока в России, составляющая около 5000 км 3 , приходится на малоплодородные и малозаселенные северные территории. При отсутствии пресной воды используют соленую поверхностную или подземную воду, производя ее опреснение или гиперфильтрацию: пропускают под большим перепадом давлений через полимерные мембраны с микроскопическими отверстиями, задерживающими молекулы соли. Оба эти процесса весьма энергоемки, поэтому представляет интерес предложение, состоящее в использовании в качестве источника пресной воды пресноводных айсбергов (или их части), которые с этой целью буксируют по воде к берегам, не имеющим пресной воды, где организуют их таяние. По предварительным расчетам разработчиков этого предложения, получение пресной воды будет примерно вдвое менее энергоемки по сравнению с опреснением и гиперфильтрацией. Важным обстоятельством, присущим водной среде, является то, что через нее в основном передаются инфекционные заболевания (примерно 80% всех заболеваний). Впрочем, некоторые из них, например, коклюш, ветрянка, туберкулез, передаются через воздушную среду. С целью борьбы с распространением заболеваний через водную среду Всемирная организация здравоохранения (ВОЗ) объявила текущее десятилетие десятилетием питьевой воды.
Водный баланс Земли
Чтобы представить, сколько воды участвует в круговороте, охарактеризуем различные части гидросферы. Более 94% ее составляет Мировой океан. Другая часть (4%) – подземные воды. При этом следует учесть, что большая их часть относится к глубинным рассолам, а пресные воды составляют 1/15 долю. Значителен также объем льда полярных ледников: с пересчетом на воду он достигает 24 млн. км., или 1,6% объема гидросферы. Озерной воды в 100 раз меньше – 230 тыс. км., а в руслах рек содержится всего лишь 1200 м. Воды, или 0,0001% всей гидросферы. Однако, несмотря на малый объем воды, реки играют очень большую роль: они, как и подземные воды, удовлетворяют значительную часть потребностей населения, промышленности и орошаемого земледелия. Воды на Земле довольно много. Гидросфера составляет около 1/4180 части массы нашей планеты. Однако на долю пресных вод, исключая воду, скованную в полярных ледниках, приходится немногим более 2 млн. км., или только 0,15% всего объема гидросферы.
Гидросфера как природная система
Гидросфера – это прерывистая водная оболочка Земли, совокупность морей, океанов, континентальных вод (включая подземные) и ледяных покровов. Моря и океаны занимают около 71% земной поверхности, в них сосредоточено около 96,5% всего объема гидросферы. Суммарная площадь всех внутренних водоемов суши составляет менее 3% ее площади. На долю ледников приходится 1,6% запасов воды в гидросферы, а их площадь составляет около 10% площади континентов.
Важнейшее свойство гидросферы – единство всех видов природных вод (Мирового океана, вод суши, водяного пара в атмосфере, подземных вод), которое осуществляется в процессе круговорота воды в природе. Движущими силами этого глобального процесса служат поступающая на поверхность Земли тепловая энергия Солнца и сила тяжести, обеспечивающие перемещение и возобновление природных вод всех видов.
Испарение с поверхности Мирового океана и с поверхности суши является начальным звеном круговорота воды в природе, обеспечивающим не только возобновление наиболее ценного его компонента – пресных воды суши, но и их высокое качество. Показателем активности водообмена природных вод служит высокая скорость их возобновления, хотя различные природные воды возобновляются (замещаются) с неодинаковой скоростью. Наиболее мобильный агент гидросферы – речные воды, период возобновления которых составляет 10-14 суток.
Преобладающая часть гидросферных вод сосредоточена в Мировом океане. Мировой океан – основное замыкающее звено круговорота воды в природе. Он отдает большую часть испаряющейся влаги в атмосферу. Водные организмы, населяющие поверхностный слой Мирового океана, обеспечивают возврат в атмосферу значительной части свободного кислорода планеты.
Огромный объем Мирового океана свидетельствует о неисчерпаемости природных ресурсов планеты. Кроме того, Мировой океан является коллектором речных вод суши, ежегодно принимая около 39 тыс. м 3 воды. Наметившееся в отдельных районах загрязнение Мирового океана грозит нарушить естественный процесс влагоооборота в его наиболее ответственном звене – испарении с поверхности океана.
Вода с точки зрения химии
Огромная роль воды в жизни человека и природы послужила причиной того, что она была одним из первых соединений, привлекших внимание ученых. Тем не менее, изучение воды еще далеко не закончено.
Общие свойства воды
Вода в силу популярности ее молекул способствует разложению контактирующих с ней молекул солей на ионы, но сама вода проявляет большую устойчивость и в химически чистой воде содержится очень мало ионов по H + и OH - .
Вода – инертный растворитель; химически не изменяется под действием большинства технических соединений, которые не растворяет. Это очень важно для всех живых организмов на нашей планете, поскольку необходимые тканям питательные вещества поступают в водных растворах в сравнительно мало измененном виде. В природных условиях вода всегда содержит то или иное количество примесей, взаимодействуя не только с твердыми и жидкими веществами, но растворяя также и газы.
Даже из свежевыпавшей дождевой воды можно выделить несколько десятков миллиграммов различных растворенных в ней веществ на каждый литр объема. Абсолютно чистую воду никогда и никому еще не удавалось получить ни в одном из ее агрегатных состояний; химически чистую воду, в значительной мере лишенную растворенных веществ, производят путем длительной и кропотливой очистки в лабораториях или на специальных промышленных установках.
В природных условиях вода не может сохранить «химическую чистоту». Постоянно соприкасаясь со всевозможными веществами, она фактически всегда представляет собой раствор различного, зачастую очень сложного свойства. В пресной воде содержание растворенных веществ обычно превышает 1 г/л. От нескольких единиц до десятков граммов на литр колеблются содержание солей в морской воде: например, в Балтийском море их всего 5 г/л, в Черном – 18, а в Красном море – даже 41 г/л.
Солевой состав морской воды в основном на 89% слагается из хлоридов (преимущественно хлорида натрия, калия, кальция), 10% приходится на сульфаты (натрия, калия, магния) и 1% - на карбонаты (натрия, кальция) и другие соли. Пресные воды содержат обычно больше всего до 80% карбонатов (натрия, кальция), около 13% сульфатов (натрия, калия, магния) и 7% хлоридов (натрия и кальция).
Вода хорошо растворяет газы (особенно при низких температурах), главным образом кислород, азот, диоксид углерода, сероводород. Количество кислорода иногда достигает 6 мг/л. В минеральных водах типа нарзан общее содержание газов может составлять до 0,1%. В природной воде присутствуют гумусовые вещества – сложные органические соединения, образующиеся в результате неполного распада остатков растительных и животных тканей, а также соединения типа белков, сахаров, спиртов.
Вода обладает исключительно высокой теплоемкостью. Теплоемкость воды принята за единицу. Теплоемкость песка, например, составляет 0,2, а железа – лишь 0,107 теплоемкости воды. Способность воды накапливать большие запасы тепловой энергии позволяет сглаживать резкие температурные колебания на прибрежных участках Земли в различные времена года и в различную пору суток: вода выступает как бы регулятором температуры на всей нашей планете.
Следует отметить особое свойство воды – ее высокое поверхностное напряжение – 72,7 эрг/см 2 (при 20°С). В этом отношении из всех видов жидкостей вода уступает только ртути. Подобное свойство воды во многом обусловлено водородными связями между отдельными молекулами H 2 O. Особенно наглядно проявляется поверхностное напряжение в прилипании воды ко многим поверхностям – смачивании. Установлено, что вещества – глина, песок, стекло, ткани, бумага и многие другие, легко смачиваемые водой, непременно имеют в своем составе атомы кислорода. Такой факт оказался ключевым при объяснении природы смачивания: энергетически неуравновешенные молекулы поверхностного слоя воды получают возможность образовать дополнительные связи с «чужими» атомами кислорода.
Смачивание и поверхностное натяжение лежат в составе явления, названного капиллярностью: в узких каналах вода способна подниматься на высоту гораздо большую, чем та, которую «позволяет» сила тяжести для столбика данного сечения.
В капиллярах вода обладает поразительными свойствами. Б. В. Дерягин установил, что в капиллярах вода, сконденсировавшаяся из водяного пара, не замерзает при 0° и даже при снижении температуры на десятки градусов.
Плотность воды - это фактор, определяющий условия передвижения водных организмов и давление на разных глубинах. Для дистиллированной воды плотность равна 1 г/см 3 при 4 °C. Плотность природных вод, содержащих растворенные соли, может быть больше, до 1,35 г/см 3 . Давление возрастает с глубиной примерно в среднем на 1 · 10 5 Па (1 атм) на каждые 10 м.
В связи с резким градиентом давления в водоемах гидробионты в целом значительно более эврибатны по сравнению с сухопутными организмами. Некоторые виды, распространенные на разных глубинах, переносят давление от нескольких до сотен атмосфер. Например, голотурии рода Elpidia, черви Priapulus caudatus обитают от прибрежной зоны до ультраабиссали. Даже пресноводные обитатели, например инфузории‑туфельки, сувойки, жуки‑плавунцы и др., выдерживают в опыте до 6 · 10 7 Па (600 атм).
Однако многие обитатели морей и океанов относительно стенобатны и приурочены к определенным глубинам. Стенобатность чаще всего свойственна мелководным и глубоководным видам. Только на литорали обитают кольчатый червь пескожил Arenicola, моллюски морские блюдечки (Patella). Многие рыбы, например из группы удильщиков, головоногие моллюски, ракообразные, погонофоры, морские звезды и др. встречаются лишь на больших глубинах при давлении не менее 4 · 10 7 - 5 · 10 7 Па (400-500 атм).
Плотность воды обеспечивает возможность опираться на нее, что особенно важно для бесскелетных форм. Плотность среды служит условием парения в воде, и многие гидробионты приспособлены именно к этому образу жизни. Взвешенные, парящие в воде организмы объединяют в особую экологическую группу гидробионтов - планктон («планктос» - парящий).
Рис. 39. Увеличение относительной поверхности тела у планктонных организмов (по С. A. Зернову, 1949):
A - палочковидные формы:
1 - диатомея Synedra;
2 - цианобактерия Aphanizomenon;
3 - перидинеевая водоросль Amphisolenia;
4 - Euglena acus;
5 - головоногий моллюск Doratopsis vermicularis;
6 - веслоногий рачок Setella;
7 - личинка Porcellana (Decapoda)

Б - расчлененные формы:
1 - моллюск Glaucus atlanticus;
2 - червь Tomopetris euchaeta;
3 - личинка рака Palinurus;
4 - личинка рыбы морского черта Lophius;
5 - веслоногий рачок Calocalanus pavo
В составе планктона - одноклеточные и колониальные водоросли, простейшие, медузы, сифонофоры, гребневики, крылоногие и киленогие моллюски, разнообразные мелкие рачки, личинки донных животных, икра и мальки рыб и многие другие (рис. 39). Планктонные организмы обладают многими сходными адаптациями, повышающими их плавучесть и препятствующими оседанию на дно. К таким приспособлениям относятся: 1) общее увеличение относительной поверхности тела за счет уменьшения размеров, сплющенности, удлинения, развития многочисленных выростов или щетинок, что увеличивает трение о воду; 2) уменьшение плотности за счет редукции скелета, накопления в теле жиров, пузырьков газа и т. п. У диатомовых водорослей запасные вещества отлагаются не в виде тяжелого крахмала, а в виде жировых капель. Ночесветка Noctiluca отличается таким обилием газовых вакуолей и капелек жира в клетке, что цитоплазма в ней имеет вид тяжей, сливающихся только вокруг ядра. Воздухоносные камеры есть и у сифонофор, ряда медуз, планктонных брюхоногих моллюсков и др.
Водоросли (фитопланктон) парят в воде пассивно, большинство же планктонных животных способно к активному плаванию, но в ограниченных пределах. Планктонные организмы не могут преодолевать течения и переносятся ими на большие расстояния. Многие виды зоопланктона способны, однако, к вертикальным миграциям в толще воды на десятки и сотни метров как за счет активного передвижения, так и за счет регулирования плавучести своего тела. Особую разновидность планктона составляет экологическая группа нейстона («нейн» - плавать) - обитатели поверхностной пленки воды на границе с воздушной средой.
Плотность и вязкость воды сильно влияют на возможность активного плавания. Животных, способных к быстрому плаванию и преодолению силы течений, объединяют в экологическую группу нектона («нектос» - плавающий). Представители нектона - рыбы, кальмары, дельфины. Быстрое движение в водной толще возможно лишь при наличии обтекаемой формы тела и сильно развитой мускулатуры. Торпедовидная форма вырабатывается у всех хороших пловцов независимо от их систематической принадлежности и способа движения в воде: реактивного, за счет изгибания тела, с помощью конечностей.
Кислородный режим. В насыщенной кислородом воде содержание его не превышает 10 мл в 1 л, это в 21 раз ниже, чем в атмосфере. Поэтому условия дыхания гидробионтов значительно усложнены. Кислород поступает в воду в основном за счет фотосинтетической деятельности водорослей и диффузии из воздуха. Поэтому верхние слои водной толщи, как правило, богаче этим газом, чем нижние. С повышением температуры и солености воды концентрация в ней кислорода понижается. В слоях, сильно заселенных животными и бактериями, может создаваться резкий дефицит О 2 из‑за усиленного его потребления. Например, в Мировом океане богатые жизнью глубины от 50 до 1000 м характеризуются резким ухудшением аэрации - она в 7‑10 раз ниже, чем в поверхностных водах, населенных фитопланктоном. Около дна водоемов условия могут быть близки к анаэробным.
Среди водных обитателей много видов, способных переносить широкие колебания содержания кислорода в воде, вплоть до почти полного его отсутствия (эвриоксибионты - «окси» - кислород, «бионт» - обитатель). К ним относятся, например, пресноводные олигохеты Tubifex tubifex, брюхоногие моллюски Viviparus viviparus. Среди рыб очень слабое насыщение воды кислородом могут выдерживать сазан, линь, караси. Вместе с тем ряд видов стеноксибионтны - они могут существовать лишь при достаточно высоком насыщении воды кислородом (радужная форель, кумжа, гольян, ресничный червь Planaria alpina, личинки поденок, веснянок и др.). Многие виды способны при недостатке кислорода впадать в неактивное состояние - аноксибиоз - и таким образом переживать неблагоприятный период.
Дыхание гидробионтов осуществляется либо через поверхность тела, либо через специализированные органы - жабры, легкие, трахеи. При этом покровы могут служить дополнительным органом дыхания. Например, рыба вьюн через кожу потребляет в среднем до 63 % кислорода. Если через покровы тела происходит газообмен, то они очень тонки. Дыхание облегчается также увеличением поверхности. Это достигается в ходе эволюции видов образованием различных выростов, уплощением, удлинением, общим уменьшением размеров тела. Некоторые виды при недостатке кислорода активно изменяют величину дыхательной поверхности. Черви Tubifex tubifex сильно вытягивают тело в длину; гидры и актинии - щупальцы; иглокожие - амбулакральные ножки. Многие сидячие и малоподвижные животные обновляют вокруг себя воду, либо создавая ее направленный ток, либо колебательными движениями способствуя ее перемешиванию. Двустворчатым моллюскам для этой цели служат реснички, выстилающие стенки мантийной полости; ракообразным - работа брюшных или грудных ножек. Пиявки, личинки комаров‑звонцов (мотыль), многие олигохеты колышут тело, высунувшись из грунта.
У некоторых видов встречается комбинирование водного и воздушного дыхания. Таковы двоякодышащие рыбы, сифонофоры дискофанты, многие легочные моллюски, ракообразные Gammarus lacustris и др. Вторичноводные животные сохраняют обычно атмосферный тип дыхания как более выгодный энергетически и нуждаются поэтому в контактах с воздушной средой, например ластоногие, китообразные, водяные жуки, личинки комаров и др.
Нехватка кислорода в воде приводит иногда к катастрофическим явлениям - заморам, сопровождающимся гибелью множества гидробионтов. Зимние заморы часто вызываются образованием на поверхности водоемов льда и прекращением контакта с воздухом; летние - повышением температуры воды и уменьшением вследствие этого растворимости кислорода.
Частая гибель рыб и многих беспозвоночных зимой характерна, например, для нижней части бассейна реки Оби, воды которой, стекающие из заболоченных пространств Западно‑Сибирской низменности, крайне бедны растворенным кислородом. Иногда заморы возникают и в морях.
Кроме недостатка кислорода, заморы могут быть вызваны повышением концентрации в воде токсичных газов - метана, сероводорода, СО 2 и др., образующихся в результате разложения органических материалов на дне водоемов.
Солевой режим. Поддержание водного баланса гидробионтов имеет свою специфику. Если для наземных животных и растений наиболее важно обеспечение организма водой в условиях ее дефицита, то для гидробионтов не менее существенно поддержание определенного количества воды в теле при ее избытке в окружающей среде. Излишнее количество воды в клетках приводит к изменению в них осмотического давления и нарушению важнейших жизненных функций.
Большинство водных обитателей пойкилосмотичны: осмотическое давление в их теле зависит от солености окружающей воды. Поэтому для гидробионтов основной способ поддерживать свой солевой баланс - это избегать местообитаний с неподходящей соленостью. Пресноводные формы не могут существовать в морях, морские - не переносят опреснения. Если соленость воды подвержена изменениям, животные перемещаются в поисках благоприятной среды. Например, при опреснении поверхностных слоев моря после сильных дождей радиолярии, морские рачки Calanus и другие спускаются на глубину до 100 м. Позвоночные животные, высшие раки, насекомые и их личинки, обитающие в воде, относятся к гомойосмотическим видам, сохраняя постоянное осмотическое давление в теле независимо от концентрации солей в воде.
У пресноводных видов соки тела гипертоничны по отношению к окружающей воде. Им угрожает излишнее обводнение, если не препятствовать поступлению или не удалять избыток воды из тела. У простейших это достигается работой выделительных вакуолей, у многоклеточных - удалением воды через выделительную систему. Некоторые инфузории каждые 2-2,5 мин выделяют количество воды, равное объему тела. На «откачку» избыточной воды клетка затрачивает очень много энергии. С повышением солености работа вакуолей замедляется. Так, у туфелек Paramecium при солености воды 2,5%о вакуоль пульсирует с интервалом в 9 с, при 5%о - 18 с, при 7,5%о - 25 с. При концентрации солей 17,5%о вакуоль перестает работать, так как разница осмотического давления между клеткой и внешней средой исчезает.
Если вода гипертонична по отношению к жидкостям тела гидробионтов, им грозит обезвоживание в результате осмотических потерь. Защита от обезвоживания достигается повышением концентрации солей также в теле гидробионтов. Обезвоживанию препятствуют непроницаемые для воды покровы гомойосмотических организмов - млекопитающих, рыб, высших раков, водных насекомых и их личинок.
Многие пойкилосмотические виды переходят к неактивному состоянию - анабиозу в результате дефицита воды в теле при возрастании солености. Это свойственно видам, обитающим в лужах морской воды и на литорали: коловраткам, жгутиковым, инфузориям, некоторым рачкам, черноморским полихетам Nereis divesicolor и др. Солевой анабиоз - средство переживать неблагоприятные периоды в условиях переменной солености воды.
Истинно эвригалинных видов, способных в активном состоянии обитать как в пресной, так и в соленой воде, среди водных обитателей не так много. В основном это виды, населяющие эстуарии рек, лиманы и другие солоноватоводные водоемы.
Температурный режим водоемов более устойчив, чем на суше. Это связано с физическими свойствами воды, прежде всего высокой удельной теплоемкостью, благодаря которой получение или отдача значительного количества тепла не вызывает слишком резких изменений температуры. Испарение воды с поверхности водоемов, при котором затрачивается около 2263,8 Дж/г, препятствует перегреванию нижних слоев, а образование льда, при котором выделяется теплота плавления (333,48 Дж/г), замедляет их охлаждение.
Амплитуда годовых колебаний температуры в верхних слоях океана не более 10-15 °C, в континентальных водоемах - 30-35 °C. Глубокие слои воды отличаются постоянством температуры. В экваториальных водах среднегодовая температура поверхностных слоев +(26-27) °С, в полярных - около 0 °C и ниже. В горячих наземных источниках температура воды может приближаться к +100 °C, а в подводных гейзерах при высоком давлении на дне океана зарегистрирована температура +380 °C.
Таким образом, в водоемах существует довольно значительное разнообразие температурных условий. Между верхними слоями воды с выраженными в них сезонными колебаниями температуры и нижними, где тепловой режим постоянен, существует зона температурного скачка, или термоклина. Термоклин резче выражен в теплых морях, где сильнее перепад температуры наружных и глубинных вод.
В связи с более устойчивым температурным режимом воды среди гидробионтов в значительно большей мере, чем среди населения суши, распространена стенотермность. Эвритермные виды встречаются в основном в мелких континентальных водоемах и на литорали морей высоких и умеренных широт, где значительны суточные и сезонные колебания температуры.
Световой режим. Света в воде гораздо меньше, чем в воздухе. Часть падающих на поверхность водоема лучей отражается в воздушную среду. Отражение тем сильнее, чем ниже положение Солнца, поэтому день под водой короче, чем на суше. Например, летний день около острова Мадейра на глубине 30 м - 5 ч, а на глубине 40 м всего 15 мин. Быстрое убывание количества света с глубиной связано с поглощением его водой. Лучи с разной длиной волны поглощаются неодинаково: красные исчезают уже недалеко от поверхности, тогда как сине‑зеленые проникают значительно глубже. Сгущающиеся с глубиной сумерки в океане имеют сначала зеленый, затем голубой, синий и сине‑фиолетовый цвет, сменяясь наконец постоянным мраком. Соответственно сменяют друг друга с глубиной зеленые, бурые и красные водоросли, специализированные на улавливании света с разной длиной волны.
Окраска животных меняется с глубиной так же закономерно. Наиболее ярко и разнообразно окрашены обитатели литоральной и сублиторальной зон. Многие глубинные организмы, подобно пещерным, не имеют пигментов. В сумеречной зоне широко распространена красная окраска, которая является дополнительной к сине‑фиолетовому свету на этих глубинах. Дополнительные по цвету лучи наиболее полно поглощаются телом. Это позволяет животным скрываться от врагов, так как их красный цвет в сине‑фиолетовых лучах зрительно воспринимается как черный. Красная окраска характерна для таких животных сумеречной зоны, как морской окунь, красный коралл, различные ракообразные и др.
У некоторых видов, обитающих у поверхности водоемов, глаза разделяются на две части с разной способностью к преломлению лучей. Одна половина глаза видит в воздухе, другая - в воде. Такая «четырехглазость» характерна для жуков‑вертячек, американской рыбки Anableps tetraphthalmus, одного из тропических видов морских собачек Dialommus fuscus. Эта рыбка при отливах сидит в углублениях, выставляя часть головы из воды (см. рис. 26).
Поглощение света тем сильнее, чем меньше прозрачность воды, которая зависит от количества взвешенных в ней частиц.
Прозрачность характеризуют предельной глубиной, на которой еще виден специально опускаемый белый диск диаметром около 20 см (диск Секки). Самые прозрачные воды - в Саргассовом море: диск виден до глубины 66,5 м. В Тихом океане диск Секки виден до 59 м, в Индийском - до 50, в мелких морях - до 5‑15 м. Прозрачность рек в среднем 1-1,5 м, а в самых мутных реках, например в среднеазиатских Амударье и Сырдарье, всего несколько сантиметров. Граница зоны фотосинтеза поэтому сильно варьирует в разных водоемах. В самых чистых водах эуфотическая зона, или зона фотосинтеза, простирается до глубин не свыше 200 м, сумеречная, или дисфотическая, зона занимает глубины до 1000-1500 м, а глубже, в афотическую зону, солнечный свет не проникает совсем.
Количество света в верхних слоях водоемов сильно меняется в зависимости от широты местности и от времени года. Длинные полярные ночи сильно ограничивают время, пригодное для фотосинтеза, в арктических и приантарктических бассейнах, а ледовый покров затрудняет доступ света зимой во все замерзающие водоемы.
В темных глубинах океана в качестве источника зрительной информации организмы используют свет, испускаемый живыми существами. Свечение живого организма получило название биолюминесценции. Светящиеся виды есть почти во всех классах водных животных от простейших до рыб, а также среди бактерий, низших растений и грибов. Биолюминесценция, по‑видимому, многократно возникала в разных группах на разных этапах эволюции.
Химия биолюминесценции сейчас довольно хорошо изучена. Реакции, используемые для генерации света, разнообразны. Но во всех случаях это окисление сложных органических соединений (люциферинов) с помощью белковых катализаторов (люцифераз). Люциферины и люциферазы у разных организмов имеют неодинаковую структуру. В ходе реакции избыточная энергия возбужденной молекулы люциферина выделяется в виде квантов света. Живые организмы испускают свет импульсами, обычно в ответ на раздражения, поступающие из внешней среды.
Свечение может и не играть особой экологической роли в жизни вида, а быть побочным результатом жизнедеятельности клеток, как, например, у бактерий или низших растений. Экологическую значимость оно получает только у животных, обладающих достаточно развитой нервной системой и органами зрения. У многих видов органы свечения приобретают очень сложное строение с системой отражателей и линз, усиливающих излучение (рис. 40). Ряд рыб и головоногих моллюсков, неспособных генерировать свет, используют симбиотических бактерий, размножающихся в специальных органах этих животных.

Рис. 40. Органы свечения водных животных (по С. А. Зернову, 1949):
1 - глубоководный удильщик с фонариком над зубатой пастью;
2 - распределение светящихся органов у рыбы сем. Mystophidae;
3 - светящийся орган рыбы Argyropelecus affinis:
а - пигмент, б - рефлектор, в - светящееся тело, г - линза
Биолюминесценция имеет в жизни животных в основном сигнальное значение. Световые сигналы могут служить для ориентации в стае, привлечения особей другого пола, подманивания жертв, для маскировки или отвлечения. Вспышка света может быть защитой от хищника, ослепляя или дезориентируя его. Например, глубоководные каракатицы, спасаясь от врага, выпускают облако светящегося секрета, тогда как виды, обитающие в освещенных водах, используют для этой цели темную жидкость. У некоторых донных червей - полихет - светящиеся органы развиваются к периоду созревания половых продуктов, причем светятся ярче самки, а глаза лучше развиты у самцов. У хищных глубоководных рыб из отряда удильщиковидных первый луч спинного плавника сдвинут к верхней челюсти и превращен в гибкое «удилище», несущее на конце червеобразную «приманку» - железу, заполненную слизью со светящимися бактериями. Регулируя приток крови к железе и, следовательно, снабжение бактерии кислородом, рыба может произвольно вызывать свечение «приманки», имитируя движения червя и подманивая добычу.
Федеральное агентство по рыболовству
ФГОУ ВПО Камчатский Государственный технический университет
Кафедра экологии и природопользования
Дисциплина экология
Реферат на тему
“Водная среда жизни и адаптация к ней организмов ”
Выполнил Проверил
Студент группы 11ПЖб Доцент
Сазонов П.А. Ступникова Н.А.
Петропавловск-Камчатский
Введение…………………………………….3
Общая характеристика……………………...3- 4
Экологические зоны Мирового океана………….4
Основные свойства водной среды………………….5
· Плотность…………………………………….5- 6
· Кислородный режим…………………………6-7
· Солевой режим……………………………….7-8
· Температурный режим………………………8
· Световой режим………………………………..8- 9
Специфические приспособления гидробионтов………..10- 11
Особенности адаптации растений к водной среде………11- 12
Особенности адаптации животных к водной среде……..12- 14
Список литературы…………………………………………15
Введение
На нашей планете живые организмы освоили четыре основные среды
обитания. Водная среда была первой, в которой возникла и
распространилась жизнь. Только потом организмы овладели
наземно-воздушной, создали и заселили почву и сами стали четвертой
специфической средой жизни.
Вода как среда обитания имеет ряд специфических свойств, таких как
большая плотность, сильные перепады давления, малое содержание
кислорода, сильное поглощение солнечных лучей. Кроме того, водоемы и
их отдельные участки различаются солевым режимом, скоростью течений,
также свойства грунта, режим разложения органических остатков и т.д.
Поэтому наряду с адаптациями к общим свойствам водной среды ее
обитатели должны быть приспособлены и к разнообразным частным
условиям.
Все обитатели водной среды получили в экологии общее название
гидробионтов.
Гидробионты населяют Мировой океан, континентальные водоемы и
подземные воды.
Общая характеристика
Гидросфера как водная среда жизни занимает около 71% площади и 1/800 часть объема земного шара. Основное количество воды, более 94%, сосредоточено в морях и океанах. В пресных водах рек, озер количество воды не превышает 0,016% общего объема пресной воды.
В океане с входящими в него морями прежде всего различают две экологические области: толщу воды - пелагиаль и дно - бенталь. В зависимости от глубины бенталь делится на сублиторальную зону - область плавного понижения суши до глубины 200 м, батиальную - область крутого склона и абиссальную зону - океанического ложа со средней глубиной 3-6 км. Более глубокие области бентали, соответствующие впадинам океанического ложа (6-10 км) называются ультраабиссалью. Кромка берега, заливаемая во время приливов, называется литоралью. Часть берега выше уровня приливов, увлажняемая брызгами прибоя, называется суперлиторалью.
Открытые воды Мирового океана также делятся на зоны по вертикали соответствующие зонам бентали: эпипелигиаль, батипелигиаль, абиссопелигиаль.
В водной среде обитает примерно 150 000 видов животных, или около 7% общего их количества и 10 000 видов растений (8%).
Удельный вес рек, озер и болот, как уже было отмечено ранее, по сравнению с морями и океанами незначителен. Однако они создают необходимый для растений, животных и человека запас пресной воды.
Характерной чертой водной среды является ее подвижность, особенно в проточных, быстро текущих ручьях и реках. В морях и океанах наблюдаются приливы и отливы, мощные течения, штормы. В озерах вода перемещается под действием температуры и ветра.
Экологические зоны Мирового океана
В любом водоеме можно выделить различные по условиям зоны. В океане
вместе с входящими в него морями различают, прежде всего, две
экологические области: пелагиаль – толща воды и бенталь –
В зависимости от глубины бенталь делится на сублиторальную зону – область плавного понижения суши до глубины
примерно 200 м, батиальную – область крутого склона и абиссальную
зону – океанического ложа со средней глубиной 3-6 км. Еще более
глубокие области бентали, соответствующие впадинам океанического ложа,
называют ультрабенталью. Кромка берега, заливаемая во время приливов,
называется литоралью. Часть берега выше уровня приливов, увлажняемая
брызгами, называется супралиторалью.
Естественно, что, например, обитатели сублиторали живут в условиях
относительно невысокого давления, дневного солнечного освещения, часто
довольно значительных изменений температурного режима. Обитатели
абиссальных и ультраабиссальных глубин существуют во мраке, при
постоянной температуре и давлении в несколько сотен, а иногда и около
тысячи атмосфер. Поэтому одно лишь указание на то, в какой зоне
бентали обитает тот или иной вид организмов, уже говорит о том, какими
общими экологическими свойствами он должен обладать.
Все население дна океана получило название бентоса. Организмы,
обитающие в толще воды, или пелагиали, относятся к пелагосу.
Пелагиаль также делят на вертикальные зоны, соответствующие по глубине
зонам бентали: эпипелагиаль, батипелагиаль, абиссопелагиаль. Нижняя
граница эпипелагиали (не более 200 м) определяется проникновением
солнечного света в количестве, достаточном для фотосинтеза. Зеленые
растения глубже этих зон существовать на могут. В сумеречных
батиальных и полных мрака абиссальных глубинах обитают лишь
микроорганизмы и животные. Разные экологические зоны выделяются и во
всех других типах водоемов: озерах, болотах, прудах, реках и т.д.
Разнообразие гидробионтов, освоивших все эти места обитания, очень
Основные свойства водной среды
1. Плотность воды
это фактор, определяющий условия передвижения водных организмов и
давление на разных глубинах. Для дистиллированной воды плотность равна
1 г/см 3 при +4 0 С. Плотность природных вод, содержащих растворенные
соли, может быть больше, до 1, 35 г/см 3 . Давление возрастает с
глубиной примерно в среднем на 1 атмосферу на каждые 10 м.
В связи с резким градиентом давления в водоемах гидробионты в целом
значительно более эврибатны по сравнению с сухопутными организмами.
Некоторые виды, распространенные на разных глубинах, переносят
давление от нескольких до сотен атмосфер.
Однако многие обитатели морей и океанов относительно стенобатны и
приурочены к определенным глубинам. Стенобатность обычно свойственна
мелководным и глубоководным видам.
Плотность воды обеспечивает возможность опираться на нее, что
особенно важно для бесскелетных форм. Опорность среды служит условием
парения в воде, и многие гидробионты приспособлены именно к этому
образу жизни. Взвешенные, парящие в воде организмы объединяют в особую
экологическую группу гидробионтов планктон.
В составе планктона одноклеточные водоросли, простейшие, медузы,
сифонофоры, гребневики, крылоногие и киленогие моллюски, разнообразные
мелкие рачки, личинки донных животных, икра и мальки рыб и многие
другие. Планктонные организмы обладают многими сходными адаптациями,
повышающими их плавучесть и препятствующими оседанию на дно. К таким
приспособлениям относятся: 1) общее увеличение поверхности тела за
счет уменьшения размеров, сплющенности, удлинения, развития
многочисленных выростов и щетинок, что увеличивает трение о воду; 2)
уменьшение плотности за счет редукции скелета, накопления в теле
жиров, пузырьков газа и т.д.
Одноклеточные водоросли фитопланктон парят в воде пассивно,
большинство же планктонных животных способно к активному плаванию, но
в ограниченных пределах. Планктонные организмы не могут преодолевать
течения и переносятся ими на большие расстояния. Многие виды
зоопланктона способны, однако, на к вертикальным миграциям в толще
воды на десятки и сотни метров как за счет активного передвижения, так
и за счет регулирования плавучести своего тела. Особую разновидность
планктона составляет экологическая группа нейстона обитатели
поверхностной пленки воды на границе с воздушной средой.
Плотность и вязкость воды сильно влияют на возможность активного
плавания. Животных, способных к быстрому плаванию и преодолению силы
течений, объединяют в экологическую группу нектона. Представители
нектона рыбы, кальмары, дельфины. Быстрое движение в водной толще
возможно, лишь при наличии обтекаемой формы тела и сильно развитой
мускулатуры. Торпедовидная форма вырабатывается у всех хороших
пловцов, независимо от их систематической принадлежности и способа
движения в воде: реактивного, за счет изгибания тела, с помощью
конечностей.
2. Кислородный режим
Коэффициент диффузии кислорода в воде примерно в 320 тыс. раз ниже,
чем в воздухе, а общее содержание его не превышает 10 мл в 1 литре
воды, это в 21 раз ниже, чем в атмосфере. Поэтому условия дыхания
гидробионтов значительно усложнены. Кислород поступает в воду в
основном за счет фотосинтетической деятельности водорослей и диффузии
из воздуха. Поэтому верхние соли водной толщи, как правило, богаче
кислородом, чем нижние. С повышением температуры и солености воды
концентрация в ней кислорода понижается. В слоях, сильно заселенных
бактериями и животными, может создаваться резкий дефицит кислорода
изза усиленного его потребления.
Среди водных обитателей много видов, способных переносить широкие
отсутствия (эвриоксибионты). Вместе с тем ряд видов стеноксибионтны
они могут существовать лишь при достаточно высоком насыщении воды
кислородом. Многие виды способны при недостатке кислорода впадать в
неактивное состояние аноксибиоз и таким образом переживать
неблагоприятный период.
Дыхание гидробионтов осуществляется либо через поверхность тела,
либо через специализированные органы жабры, легкие, трахеи.
При этом покровы могут служить дополнительным органом дыхания. Если
через покровы тела происходит газообмен, то они очень тонки. Дыхание
облегчается также увеличением поверхности. Это достигается в ходе
эволюции видов образованием различных выростов, уплощением,
удлинением, общим уменьшением размеров тела. Некоторые виды при
нехватке кислорода активно изменяют величину дыхательной поверхности.
Многие сидячие и малоподвижные животные обновляют вокруг себя воду,
либо создавая ее направленный ток, либо колебательными движениями
способствуя ее перемешиванию.
У некоторых видов встречается комбинирование водного и воздушного
дыхания. Вторичноводные животные сохраняют обычно атмосферное дыхание
как более выгодный энергетически и нуждаются поэтому в контактах с
воздушной средой.
Нехватка кислорода в воде приводит иногда к катастрофическим
явлениям заморам, сопровождающимся гибелью множества гидробионтов.
Зимние заморы часто вызываются образованием на поверхности водоемов
льда и прекращением контакта с воздухом; летние повышением температуры
воды и уменьшением вследствие этого растворимости кислорода. Заморы
чаще возникают чаще возникают в прудах, озерах, реках. Реже заморы
происходят в морях. Кроме недостатка кислорода, заморы могут быть
вызваны повышением концентрации в воде токсичных газов метана,
сероводорода и других, образующихся в результате разложения
органических материалов на дне водоемов.
3. Солевой режим
Поддержание водного баланса гидробионтов имеет свою специфику. Если
для наземных животных и растений наиболее важно обеспечение организма
водой в условиях ее дефицита, то для гидробионтов не менее существенно
поддержание определенного количества воды в теле при ее избытке в
окружающей среде. Излишнее количество воды в клетках приводит к
изменению в них осмотического давления и нарушению важнейших жизненных
Большинство водных обитателей пойкилосмотичны: осмотическое давление
в их теле зависит от солености окружающей воды. Поэтому для
гидробионтов основной способ поддерживать свой солевой баланс это
избегать местообитаний с неподходящей соленостью. Пресноводные формы
не могут существовать в морях, морские не переносят опреснения. Если
соленость воды подвержена изменениям, животные перемещаются в поисках
благоприятной среды. Позвоночные животные, высшие раки, насекомые и их
личинки, обитающие в воде, относятся к гомойосмотическим видам,
сохраняя постоянное осмотическое давление в теле независимо от
концентрации солей в воде.
У пресноводных видов соки тела гипертоничны по отношению к
окружающей среде. Им угрожает излишнее обводнение, если не
препятствовать поступлению или не удалять избыток воды из тела. У
простейших это достигается работой выделительных вакуолей, у
многоклеточных удалением воды через выделительную систему. Некоторые
инфузории каждые 2-2,5 минуты выделяют количество воды, равное объему
тела. На «откачку» избыточной воды клетка затрачивает очень много
энергии. С повышением солености работа вакуолей замедляется.
Если вода гипертонична по отношению к сокам тела гидробионтов, им
грозит обезвоживание в результате осмотических потерь. Защита от
обезвоживания достигается повышением концентрации солей также в теле
гидробионтов. Обезвоживанию препятствуют непроницаемые для воды
покровы гомойосматических организмов млекопитающих, рыб, высших раков,
водных насекомых и их личинок. Многие пойкилосмотические виды
переходят к неактивному состоянию анабиозу в результате дефицита воды
в теле при возрастании солености. Это свойственно видам, обитающим в
лужах морской воды и на литорали: коловраткам, жгутиковым, инфузориям,
некоторым рачкам и др. Солевой анабиоз средство переживать
неблагоприятные периоды в условиях переменной солености воды.
Истинно эвригалинных видов, способных в активном состоянии обитать
как в пресной, так и в соленой воде, среди водных обитателей не так уж
много. В основном это виды, населяющие эстуарии рек, лиманы и другие
солоноватоводные водоемы.
4. Температурный режим водоемов
более устойчив, чем на суше. Это связано с физическими свойствами
воды, прежде всего высокой удельной теплоемкостью, благодаря которой
получение или отдача значительного количества тепла не вызывает
слишком резких изменений температуры. Амплитуда годовых колебаний
температуры в верхних слоях океана не более 10-15 0 С, в
континентальных водоемах 30-35 0 С. Глубокие слои воды отличаются
постоянством температуры. В экваториальных водах среднегодовая
температура поверхностных слоев +26...+27 0 С, в полярных около 0 0 С
и ниже. Таким образом, в водоемах существует довольно значительное
разнообразие температурных условий. Между верхними слоями воды с
выраженными в них сезонными колебаниями температуры и нижними, где
тепловой режим постоянен, существует зона температурного скачка, или
термоклина. Термоклин резче выражен в теплых морях, где сильнее
перепад температуры наружных и глубинных вод.
В связи с более устойчивым температурным режимом воды среди
гидробионтов в значительно большей мере, чем среди населения суши,
распространена стенотермность. Эвритермные виды встречаются в основном
в мелких континентальных водоемах и на литорали морей высоких и
умеренных широт, где значительны суточные и сезонные колебания
температуры.
5. Световой режим водоемов
Света в воде гораздо меньше, чем в воздухе. Часть падающих на
поверхность водоема лучей отражается в воздушную среду. Отражение тем
сильнее, чем ниже положение Солнца, поэтому день под водой короче, чем
на суше. Быстрое убывание количества света с глубиной связано с
поглощением его водой. Лучи с разной длиной волны поглощаются
неодинаково: красные исчезают уже недалеко от поверхности, тогда как
синезеленые проникают гораздо глубже. Сгущающиеся с глубиной сумерки
имеют сначала зеленый, затем голубой, синий и синефиолетовый цвет,
сменяясь наконец постоянным мраком. Соответственно сменяют друг друга
с глубиной зеленые, бурые и красные водоросли, специализированные на
улавливание света с разной длиной волны. Окраска животных меняется с глубиной так же закономерно.
Наиболее ярко и разнообразно окрашены обитатели литоральной и
сублиторальной зон. Многие глубинные организмы, подобно пещерным, не
имеют пигментов. В сумеречной зоне широко распространена красная
окраска, которая является дополнительной к синефиолетовому свету на
этих глубинах. Дополнительные по цвету лучи наиболее полно поглощаются
телом. Это позволяет животным скрываться от врагов, так как их красный
цвет в синефиолетовых лучах зрительно воспринимается как черный.
Поглощение света тем сильнее, чем меньше прозрачность воды, которая
зависит от количества взвешенных в ней частиц. Прозрачность
характеризуют предельной глубиной, на которой еще виден специально
опускаемый белый диск диаметром около 20 см (диск Секки).
Специфические приспособления гидробионтов
Способы ориентации животных в водной среде
Жизнь в постоянных сумерках или во мраке сильно ограничивает
возможности зрительной ориентации гидробионтов. В связи с быстрым
затуханием световых лучей в воде даже обладатели хорошо развитых
органов зрения ориентируются при их помощи лишь на близком расстоянии.
Звук распространяется в воде быстрее, чем в воздухе. Ориентация на
звук развита у гидробионтов в целом лучше, чем зрительная. Ряд видов
улавливает даже колебания очень низкой частоты (инфразвуки),
возникающие при изменении ритма волн, и заблаговременно спускается
перед штормом из поверхностных слоев в более глубокие. Многие
обитатели водоемов млекопитающие, рыбы, моллюски, ракообразные сами
издают звуки. Ракообразные осуществляют это трением друг о друга
разных частей тела; рыбы с помощью плавательного пузыря, глоточных
зубов, челюстей, лучей грудных плавников и другими способами. Звуковая
сигнализация служит чаще всего для внутривидовых взаимоотношений
например, для ориентации в стае, привлечения особей другого пола, и
особенно развита у обитателей мутных вод и больших глубин, живущих в
Ряд гидробионтов отыскивает пище и ориентируется при помощи
эхолокации восприятия отраженных звуковых волн. Многие воспринимают
отраженные электрические импульсы, производя при плавании разряды
разной частоты. Известно около 300 видов рыб, способных генерировать
электричество и использовать его для ориентации и сигнализации. Ряд
рыб использует электрические поля также для защиты и нападения.
Для ориентации в глубине служит восприятие гидростатического давления. Оно осуществляется при помощи статоцистов, газовых камер и
других органов.
Наиболее древний способ, свойственный всем водным животным,
восприятие химизма среды. Хеморецепторы многих гидробионтов обладают
чрезвычайной чувствительностью. В тысячекилометровых миграциях,
которые характерны для многих видов рыб, они ориентируются в основном
по запахам, с поразительной точностью находя места нерестилищ или
Фильтрация как тип питания
Некоторые гидробионты обладают особым характером питания это
отцеживание или осаждение взвешенных в воде частиц органического
происхождения и многочисленных мелких организмов. Такой способ
питания, не требующий больших затрат энергии на поиски добычи,
характерен для пластинчатожабренных моллюсков, сидячих иглокожих,
полихет, мшанок, асцидий, планктонных рачков и других. Животные
фильтраторы выполняют важнейшую роль в биологической очистке водоемов.
Литоральная зона океана, особенно богатая скоплениями фильтрующих
организмов, работает как эффективная очистительная система.
Специфика приспособлений к жизни в пересыхающих водоемах
На Земле существует много временных, неглубоких водоемов,
возникающие после разлива рек, сильных дождей, таяния снега и т.п. В
этих водоемах, несмотря на краткость их существования, поселяются
разнообразные гидробионты. Общими особенностями обитателей
пересыхающих бассейнов являются способности давать за короткие сроки
многочисленное потомство и переносить длительные периоды без воды.
Представители многих видов при этом закапываются в ил, переходя в
состояние пониженной жизнедеятельности гипобиоза. Многие мелкие виды
образуют цисты, выдерживающие засуху. Другие переживают
неблагоприятный период в стадии высокоустойчивых яиц. Некоторым видам
пересыхающих водоемов присуща уникальная способность высыхать до
состояния пленки, а при увлажнении возобновлять рост и развитие.
Экологическая пластичность является важным регулятором расселения организмов. Гидробионты с высокой экологической пластичностью распространены широко, например, элодея. Противоположный пример - рачок артемия, живущий в небольших водоемах с очень соленой водой, является типичным стеногалинным представителем с узкой экологической пластичностью. По отношению же к другим факторам он обладает значительной пластичностью и в соленых водоемах встречается довольно часто.
Экологическая пластичность зависит от возраста и фазы развития организма. Например, морской брюхоногий моллюск Littorina во взрослом состоянии при отливах ежедневно длительное время находится без воды, однако его личинки ведут планктонный образ жизни и не переносят высыхания.
Особенности адаптации растений к водной среде
Водные растения имеют значительные отличия от наземных растительных организмов. Так, способность водных растений поглощать влагу и минеральные соли непосредственно из окружающей среды отражается на их морфологической и физиологической организации. Характерным для водных растений является слабое развитие проводящей ткани и корневой системы. Корневая система служит главным образом для прикрепления к подводному субстрату и не выполняет функции минерального питания и водоснабжения, как у наземных растений. Питание же водных растений осуществляется всей поверхностью их тела. Значительная плотность воды дает возможность обитания растений во всей ее толще. У низших растений, заселяющих различные слои и ведущих плавающий образ жизни, для этого имеются специальные придатки, которые увеличивают их плавучесть и позволяют им удерживаться во взвешенном состоянии. Высшие гидрофиты имеют слабо развитую механическую ткань. В их листьях, стеблях, корнях располагаются воздухоносные межклеточные полости, увеличивающие легкость и плавучесть взвешенных в воде и плавающих на поверхности органов, что также способствует омыванию внутренних клеток водой с растворенными в ней солями и газами. Гидрофиты отличаются большой поверхностью листьев при малом общем объеме растения, что обеспечивает им интенсивный газообмен при недостатке растворенного в воде кислорода и других газов.
У ряда водных организмов развита разнолистность, или гетерофилия. Так, у сальвинии погруженные листья обеспечивают минеральное питание, а плавающие - органическое.
Важной особенностью адаптации растений к обитанию в водной среде является и то, что листья, погруженные в воду, как правило, очень тонкие. Часто хлорофилл в них располагается в клетках эпидермиса, что способствует усилению интенсивности фотосинтеза при слабом освещении. Такие анатомо-морфологические особенности наиболее четко выражены у водных мхов, валиснерии, рдестов.
От вымывания у водных растений из клеток минеральных солей или выщелачивания защитой является выделение специальными клетками слизи и образование эндодермы из более толстостенных клеток в виде кольца.
Относительно низкая температура водной среды обусловливает отмирание вегетирующих частей у погруженных в воду растений после образования зимних почек и замену летних тонких нижних листьев более жесткими и короткими зимними. Низкая температура воды отрицательно сказывается на генеративных органах водных растений, а высокая ее плотность затрудняет перенос пыльцы. В связи с этим водные растения интенсивно размножаются вегетативным путем. Большинство плавающих на поверхности и погруженных растений выносят цветоносные стебли в воздушную среду и размножаются половым путем. Пыльца разносится ветром и поверхностными течениями. Плоды и семена, которые образуются, также распространяются поверхностными течениями. Это явление носит название гидрохории. К гидрохорным относятся не только водные, а также многие прибрежные растения. Их плоды имеют высокую плавучесть, длительное время находятся в воде и не теряют при этом всхожесть. Например, водой переносятся плоды и семена стрелолиста, сусака, частухи. Плоды многих осок заключены в своеобразные мешочки с воздухом и разносятся водными течениями.
Особенности адаптации животных к водной среде
У животных, обитающих в водной среде, по сравнению с растениями адаптивные особенности более многообразны, к ним относятся такие, как анатомо-морфологические, поведенческие и др.
Животные, обитающие в толще воды, обладают в первую очередь приспособлениями, которые увеличивают их плавучесть и позволяют противостоять движению воды, течениям. Данные организмы вырабатывают приспособления, которые препятствуют поднятию их в толщу воды или уменьшают плавучесть, что позволяет удерживаться на дне, включая и быстро текущие воды.
У мелких форм, живущих в толще воды, отмечается редукция скелетных образований. Так, у простейших (радиолярии) раковины обладают пористостью, кремневые иглы скелета внутри полые. Удельная плотность гребневиков, медуз уменьшается благодаря наличию воды в тканях. Скопление капелек жира в теле способствует увеличению плавучести. Крупные скопления жира наблюдаются у некоторых ракообразных, рыб и китообразных. Удельную плотность тела снижают и тем самым повышают плавучесть плавательные пузыри, наполненные газом, которые имеют многие рыбы. У сифонофор развиты мощные воздухоносные полости.
Для животных, пассивно плавающих в толще воды, характерно не только уменьшение массы, но и увеличение удельной поверхности тела. Это связано с тем, что чем больше вязкость среды и выше удельная поверхность тела организма, тем он медленнее погружается в воду. У животных уплощается тело, на нем образуются шипы, выросты, придатки, например у жгутиковых, радиолярий.
Большая группа животных, обитающих в пресной воде, при передвижении использует поверхностное натяжение воды. По поверхности воды свободно бегают клопы водомерки, жуки вертячки и др. Членистоногое, касающееся воды окончанием своих придатков, покрытых водоотталкивающими волосками, вызывает деформацию ее поверхности с образованием вогнутого мениска. Когда подъемная сила, направленная вверх, больше массы животного, последнее и будет удерживаться на воде благодаря поверхностному натяжению.
Таким образом, жизнь на поверхности воды возможна для сравнительно мелких животных, так как масса растет пропорционально кубу размера, а поверхностное натяжение увеличивается как линейная величина.
Активное плавание у животных осуществляется с помощью ресничек, жгутиков, изгибания тела, реактивным способом за счет энергии выбрасываемой струи воды. Наибольшего совершенства реактивный способ передвижения достиг у головоногих моллюсков.
У крупных животных нередко имеются специализированные конечности (плавники, ласты), тело их обтекаемой формы и покрыто слизью.
Только в водной среде встречаются неподвижные, ведущие прикрепленный образ жизни, животные. Это такие, как гидроиды и коралловые полипы, морские лилии, двустворчатые и др. Для них характерны своеобразная форма тела, незначительная плавучесть (плотность тела больше плотности воды) и специальные приспособления для прикрепления к субстрату.
Водные животные большей частью пойкилотермны. У гомойотермных же (китообразные, ластоногие) образуется значительный слой подкожного жира, который выполняет теплоизоляционную функцию.
Глубоководные животные отличаются специфическими чертами организации: исчезновение или слабое развитие известкового скелета, увеличение размеров тела, нередко - редукция органов зрения, усиление развития осязательных рецепторов и т.д.
Осмотическое давление и ионное состояние растворов в теле животных обеспечивается сложными механизмами водно-солевого обмена. Наиболее распространенным способом поддержания постоянного осмотического давления является регулярное удаление поступающей в организм воды с помощью пульсирующих вакуолей и органов выделения. Так, пресноводные рыбы избыток воды удаляют усиленной работой выделительной системы, а соли поглощают через жаберные лепестки. Морские рыбы вынуждены пополнять запасы воды и поэтому пьют морскую воду, а излишки поступающих с водой солей выводят из организма через жаберные лепестки.
Целый ряд гидробионтов обладают особым характером питания - это отцеживание или осаждение взвешенных в воде частиц органического происхождения, многочисленных мелких организмов. Этот способ питания не требует больших затрат энергии на поиски добычи и характерен для пластинчатожаберных моллюсков, сидячих иглокожих, асцидий, планктонных рачков и др. Животные-фильтраторы выполняют важную роль в биологической очистке водоемов.
В связи с быстрым затуханием световых лучей в воде жизнь в постоянных сумерках или во мраке сильно ограничивает возможности зрительной ориентации гидробионтов. Звук распространяется в воде быстрее, чем в воздухе, и ориентация на звук у гидробионтов развита лучше зрительной. Отдельные виды улавливают даже ультразвуки. Звуковая сигнализация служит больше всего для внутривидовых взаимоотношений: ориентации в стае, привлечения особей другого пола и т.д. Китообразные, например, отыскивают пищу и ориентируются при помощи эхолокации - восприятия отраженных звуковых волн. Принцип локатора дельфина заключается в излучении звуковых волн, которые распространяются перед плывущим животным. Встречая препятствие, например рыбу, звуковые волны отражаются и возвращаются к дельфину, который слышит возникающее эхо и таким образом обнаруживает предмет, вызывающий отражение звука.
Известно около 300 видов рыб, которые способны генерировать электричество и использовать его для ориентации и сигнализации. Ряд рыб (электрический скат, электрический угорь) используют электрические поля для защиты и нападения.
Водным организмам свойственен древний способ ориентации - восприятие химизма среды. Хеморецепторы многих гидробионтов (лососи, угри) обладают чрезвычайной чувствительностью. В тысячекилометровых миграциях они с поразительной точностью находят места нерестилищ и нагула.
Список литературы
1. Акимова Т.А. Экология / Т.А. Акимова, В.В. Хаскин М.: ЮНИТИ, 1998 г.
2. Одум Ю. Общая экология / Ю. Одум М.: Мир. 1986 г.
3. Степановских А.С. Экология / А.С. Степановских М.: ЮНИТИ - 2001
4. Экологический энциклопедический словарь. М.: "Ноосфера", 1999 г.
| Наименование параметра | Значение |
| Тема статьи: | Водная среда. |
| Рубрика (тематическая категория) | Экология |
Вода является первой средой жизни: в ней возникла жизнь и сформировались большинство групп организмов. Все обитатели водной среды носят название гидробионты. Характерной чертой водной сред является движение воды, ĸᴏᴛᴏᴩᴏᴇ проявляется в форме течений (перенос воды в одном направлении) и волнений (уклонение частиц воды от исходного положения с последующим возвратом к нему). Гольфстрим переносит в год 2,5 млн м^3 воды, что в 25 раз больше всех рек Земли вместе взятых. Кроме того происходят приливно-отливные колебания уровня моря под действием притяжения Луны и Солнце.
Кроме движения воды к числу важных свойств водной среды относятся плотность и вязкость, призрачность, растворенный кислород и содержание минеральных веществ.
Плотность и вязкость определяют, в первую очередь, условия передвижения гидробионтов. Чем выше плотность вод, тем она становится более опорной, тем легче удержаться в ней. Другое значение плотности - ее давление на организм. С углублением на 10,3 м в пресную и на 9,986 м в морскую воду давление возрастает на 1 атм. С повышением вязкости возрастает сопротивление активному движению организмов. Плотность живых тканей выше, чем плотность пресной и морской воды, в связи с этим у водных организмов в процессе эволюции выработались различные структуры, которые повышают их плавучесть -общее увеличение относительной поверхности тела за счёт уменьшения размеров; сплющенность; развитие различных выростов (щетинок); уменьшение плотности тела за счёт редукции скелета; накопление жира и наличеи плавательного пузыря. Вода, в отличие от воздуха, обладает большей выталкивающей силой, в связи с этим максимальные размеры водных организмов ограничены меньше.
Термические свойства воды существенно отличаются от термических свойств воздух. Высокие удельная теплоемкость воды (в 500 раз выше) и теплопроводность (в 30 раз) определяют постоянное и относительно равномерное распределение температуры в водной среде. Колебания температуры в воде не столь резкие, как в воздухе. Температура оказывает влияние на скорость протекания различных процессов.
Свет и световой режим. Солнце с одинаковой интенсивностью освещает поверхности суши и океана, однако у воды способность к поглощению и рассеиванию достаточно велика, что ограничивает глубину проникновения света в толщу океана. Причем лучи с разной длиной волны поглощаются не одинаково: красный рассеивается почти сразу, а синие и зеленые уходят глубже. Зона в которой интенсивность фотосинтеза превосходит интенсивность дыхания принято называть эвфотической зоной. Нижняя граница на которой фотосинтез уравновешивается дыханием принято называть компенсационной точкой.
Прозрачность воды зависит от содержания в ней взвешенных частиц. Прозрачность характеризуется предельной глубиной на которой еще виден специально опускаемый белый диск диаметром 30 см. Самые прозрачные воды в Саргассовом море (диск виден на глубине 66 м), в Тихом океане (60 м), Индийском океане (50 м). В мелких морях прозрачность 2-15 м, в реках 1-1,5 м.
Кислород - необходим для дыхания. В воде распределение растворенного кислорода подвержено резким колебаниям. В ночное время содержание кислорода в воде меньше. Дыхание гидробионтов осуществляется либо через поверхность тела, либо посредством специальных органов (легки, жабры, трахея).
Минеральные вещества. Морская вода в основном содержит ионы натрия, магния, хлора, сульфат-ион. Пресная ионы кальция и карбонат-ион.
Экологическая классификация водных организмов . В воде обитают более 150 тыс. видов животных и около 10 тыс. видов растений. Основными биотопами гидробионтов являются: толща воды (пелагиаль ) и дно водоемов (бенталь ). Различают пелагиальные организмы и бентосные. Пелагиаль делится на группы: планктон (совокупность организмов, которые не способны к активному движении и перемещаются с потоками воды) и нектон (крупные животные, двигательной активности которых достаточно для преодоления водных течений). Бентос - совокупность организмов, населяющих дно.
Водная среда. - понятие и виды. Классификация и особенности категории "Водная среда." 2017, 2018.
Среда обитания, условия и образ жизни Практическое применение палеонтологии в геологии § В стратиграфии (в основе использования лежит закон необратимости эволюции). § В палеогеографииТрофические, или пищевые, связи (греч. trophe - пища, питание) – главные в... .
Беспроводная среда не означает полное отсутствие проводов в сети. Обычно беспроводные компоненты взаимодействуют с сетью, в которой в качестве среды передачи используется кабель. Такие сети называются гибридными. Существуют следующие типы беспроводных сетей: ЛВС,...
-
Экологическая система (экосистема) – пространственно определенная совокупность, состоящая из сообщества живых организмов (биоценоз), среды их обитания (биотоп), системы связей, осуществляющей обмен веществом и энергией между ними. Различают водные и наземные природные... .