,
рептилии (пресмыкающиеся) ,
птицы, их гнезда, яйца и голоса , и
млекопитающие (звери) и следы их жизнедеятельности ,
20
цветных ламинированных определительных таблиц
, в том числе:
водные беспозвоночные ,
дневные бабочки ,
рыбы ,
амфибии и рептилии ,
зимующие птицы ,
перелетные птицы ,
млекопитающие и их следы ,
4
карманных полевых определителя
,
в том числе:
обитатели водоемов ,
птицы средней полосы и
звери и их следы , а также
65
методических пособий
и
40
учебно-методических фильмов
по методикам
проведения научно-исследовательских работ в природе (в полевых условиях).
Анатомия амфибий: общий обзор
Строение тела или анатомия.
Тело
разделено на голову, туловище, хвост
(только у хвостатых и безногих) и конечности,
которые могут и отсутствовать. Голова
подвижна, соединена с туловищем. Скелет и
позвоночник разделён на отделы
. К туловищным
позвонкам прикрепляются рёбра, если они есть.
Земноводные имеют две пары первичнопятипалых конечностей ; зачаточные формы шейной части обеспечивают им возможность самостоятельного движения головой.

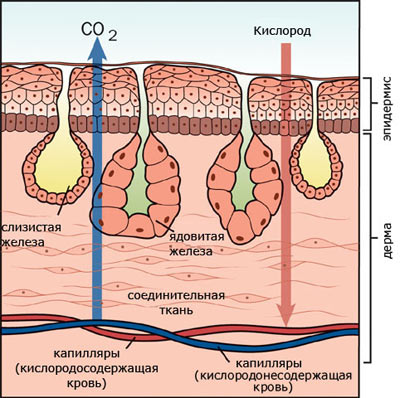
Кожа голая, лишенная чешуи. Эпидермис богат многоклеточными железами, которые обеспечивают наличие на поверхности кожи жидкостной пленки, без которой при кожном дыхании невозможен газообмен. Эпидермис многослойный, кориум тонкий, но обильно насыщенный капиллярами.
В нижних слоях эпидермиса и в кориуме расположены пигментные клетки , обусловливающие видоспецифичность окраски.
Скелет конечностей образован скелетом пояса конечностей и скелетом свободных конечностей.
Плечевой пояс лежит в толще мускулатуры и включает парные лопатки, ключицы и вороньи кости, соединённые с грудиной. Скелет передней конечности состоит из плеча (плечевая кость), предплечья (лучевая и локтевая кости) и кисти (кости запятстья, пястья и фаланги пальцев).
Тазовый пояс состоит из парных подвздошных седалишных и лобковых костей, сросшихся между собой. Он прикреплен к крестцовому позвонку через подвздошные кости. В состав скелета задней конечности входят бедро, голень (большая и малая берцовая кости) и стопа. Кости предплюсны, плюсны и фаланги пальцев. Крестец состоит из одного только позвонка.
Двигательная система.
Характер движения земноводных довольно однообразен
и может быть сведен к двум основным типам.
Ископаемые и современные хвостатые земноводные сохранили свойственный рыбам основной тип движения - при помощи сильных боковых изгибов всего тела, но с опорой на короткие ножки при движении по земле. При коротких конечностях боковые изгибы туловища увеличивают длину шага, а изгибы хвоста помогают сохранять равновесие. При движении в воде конечности сколько-нибудь заметной роли не играют. С помощью изгибов всего тела передвигаются и безногие.
Бесхвостые земноводные по суше передвигаются прыжками , поднимая тело в воздух резким толчком обоих задних конечностей. Коротконогие виды, например жабы , помимо прыжков могут медленно шагать , последовательно переставляя конечности.
В воде бесхвостые плавают , энергично работая задними конечностями (стиль "брасс", но без участия передних конечностей). Предполагают, что мощные задние конечности развились в результате приспособления к плаванию, и лишь позднее использованы для прыжков на суше.
У амфибий довольно крупная, широкая голова , которая переходит непосредственно в широкое и короткое тело . Лобные и теменные кости слиты в парную лобнотеменную кость. В черепе характерно то, что челюстно-нёбный аппарат и квадратная кость соединены с черепом неподвижно; два мыщелка черепа принадлежат собственно слившемуся с ним первому шейному позвонку, так что первый позвонок амфибий есть в сущности второй.
Мозг
амфибий отличается от мозга рыб
большим развитием переднего отдела (передний
мозг
), содержащего большое число нервных
клеток (серое вещество). Полушария
переднего
мозга малы и полностью разделены. Отделы мозга
лежат в одной горизонтальной плоскости. Обонятельные
доли сильно развиты. Мозжечок
развит очень
слабо в связи с малой подвижностью и
однообразным характером движений.
Черепномозговых нервов 10 пар. У личинок имеются
органы боковой линии
.
Спинной мозг
развит лучше головного.
Головной мозг состоит из 5 отделов
: передний
мозг, промежуточный, продолговатый, средний,
мозжечок. Промежуточный
мозг хорошо развит. Продолговатый
мозг является центром дыхательной, кровеносной и
пищеварительной системы. Средний
мозг
относительно невелик.
Органы осязания хорошо развиты. Органы боковой линии сигнализируют амфибиям о волнообразных колебаниях воды. Они даны им для активной локации водного пространства, особенно в мутной воде или в ночное время, и полностью заменяют зрение. Являясь органами дистанционного осязания, такие живые приборы чувствуют и колебания, вызванные движениями подводных жителей. Органы боковых линий находятся на поверхности кожи земноводных, живущих исключительно в воде, и для каждого вида имеются свои особенности.
Органом осязания служит вся кожа , в которой имеются осязательные нервные окончания.
Во рту также находятся органы осязания в виде вкусовых луковичек . Зубы могут как присутствовать у некоторых видов, так и отсутствовать. Зубы, как и у пресмыкающихся, приспособлены лишь к схватыванию и к удерживанию добычи, но не могут служить для ее разжевывания. Звуки могут издавать только бесхвостые амфибии , да и то преимущественно самцы.
Носовая полость снабжена задними носовыми отверстиями и носослезными каналами.
Глаза похожи на глаза рыб, но не имеют серебристой оболочки, ни отражательной, ни серповидного отростка. Аккомодация глаза осуществляется перемещением хрусталика. Глаза приспособлены к видению на большом расстоянии. Слезные железы отсутствуют, но есть Гардерова железа, секрет которой смачивает роговицу и предохраняет её от высыхания. Роговица выпуклая. Хрусталик имеет форму двояковыпуклой линзы, диаметр которой меняться в зависимости от освещения; аккомодация происходит засчёт изменения расстояния хрусталика до сетчатки. У многих развито цветное зрение .

Строение уха различается у бесхвостых и хвостатых амфибий.
Мускулатура подразделяется на мускулатуру туловища и конечностей. Туловищная мускулатура сегментирована. Группы специальных мышц обеспечивают сложные движения рычажных конечностей. На голове расположены мышцы поднимающие и опускающие. Благодаря сокращениям мышц или групп мышц амфибии могут совершать сложные движения. Особенно хорошо развиты мышцы конечностей.
Пищеварительная система земноводных имеет почти такое же строение, как у рыб. Все земноводные питаются только подвижной добычей . На дне ротоглоточной полости находятся язык. В ротоглоточную полость открываются протоки слюнных желез, секрет которых не содержит пищеварительных ферментов. Из ротоглоточной полости пища по пищеводу поступает в желудок, оттуда в двенадцатипёрстную кишку. Сюда открываются протоки печени и поджелудочной железы. Переваривание пищи происходит в желудке и в двенадцатипёрстной кишке. Тонкий кишечник переходит в толстый, заканчивающийся прямой кишкой, которая образует расширение - клоаку. В отличие от рыб задняя кишка открывается не прямо наружу, а в особое ее расширение, называемое клоакой. В клоаку открываются также мочеточники и выводные протоки органов размножения.
Органами дыхания у земноводных являются:
- лёгкие (специальные органы воздушного дыхания);
- кожа и слизистая выстилка ротоглоточной полости (дополнительные органы дыхания);
- жабры (у некоторых водных обитателей и у головастиков).
У большинства видов (кроме безлёгочных саламандр) имеются лёгкие небольшого объёма, в виде тонкостенных мешков, оплетённых густой сетью кровеносных сосудов. Каждое лёгкое открывается самостоятельным отверстием в гортанно-трахейную впадину (здесь расположены голосовые связки, открывающиеся щелью в ротоглоточную полость). Воздух нагнетается в лёгкие за счёт изменения объема ротоглоточной полости: воздух поступает в ротоглоточную полость через ноздри при опускании её дна. При поднимании дна воздух проталкивается в лёгкие.
Горло несколько раз в секунду оттягивается вниз , благодаря чему в ротовой полости создается разреженное пространство. Тогда воздух проникает через ноздри в ротовую полость, а оттуда в легкие. Обратно он выталкивается под действием мускулатуры стенок тела. Погружённая в воду амфибия целиком переходит на кожное дыхание .
Кровеносная система замкнутая, состоит из большого и малого кругов кровообращения. Появление второго круга связано с приобретением лёгочного дыхания. В организме есть кожнолёгочные артерии (несут венозную кровь к лёгким и коже), сонные артерии (снабжены артериальной кровью органы головы), дуги аорты несут смешанную кровь к остальным органам тела.

I - венозная пазуха; II - правое предсердие; III - левое предсердие; IV - желудочек; V - артериальный ствол;
1 - лёгочно-кожная артерия; 2 - дуга аорты; 3 - сонная артерия; 4 - язычная артерия; 5 - сонная железа; 6 - подключичная артерия; 7 - общая аорта; 8 - кишечная артерия; 9 - кожная артерия; 10 - лёгочная вена; 11 - лёгкое; 12 - задняя полая вена; 13 - кожная вена; 14 - брюшная вена; 15 - печень; 16 - почечная вена.
Малый круг кровообращения - лёгочный, начинается кожно-лёгочными артериями, несущими кровь к органам дыхания (лёгким и коже); от лёгких обогащённая кислородом кровь собирается в парные лёгочные вены, впадающие в левое предсердие.
Большой круг кровообращения начинается дугами аорты и сонными артериями, которые ветвятся в органах и тканях. Венозная кровь по парным передним полым венам и непарной задней полой вене попадает в правое предсердие. Кроме того, в передние полые вены попадает окисленная кровь, и поэтому кровь в правом предсердии смешанная. Так как органы тела снабжаются смешанной кровью, у амфибий низкий уровень обмена веществ и поэтому они хладнокровные животные.
Аорта проходит в жаберные дуги и разветвляется сначала в наружных жабрах, а позднее и во внутренних. Обратно кровь течет по вене, идущей вдоль хвоста, а затем разветвляется на поверхности желточного пузыря и через желточные вены возвращается обратно в предсердие. Позднее постепенно образуются воротные системы печени и почек. В конце личиночной стадии жаберное дыхание мало-помалу заменяется легочным; передние жаберные дуги превращаются в головные артерии, а средние образуют аорту.
Сердце трёхкамерное. Состоит из двух предсердий (в правом предсердии кровь смешанная, преимущественно венозная, а в левом - артериальная) и одного желудочка. Внутри стенки желудочка образуют складки, препятствующие смешиванию артериальной и венозной крови. Из желудочка выходит артериальный конус, снабжённый спиральным клапаном.
Правое предсердие получает венозную кровь, левое - артериальную (из лёгких и кожи). Венозная и артериальная кровь лишь частично смешиваются в полости желудочка, стенки которого имеют сложную систему мускульных перекладин. В лёгочные вены направляется главным образом венозная кровь, дуги аорты заполняются смешанной кровью, и лишь сонные артерии получают артериальную кровь.
Сердце образуется у личинок очень рано и тотчас же начинает действовать. Первоначально оно представляет простой мешок, который впоследствии разделяется на отдельные части.
Органы выделения - парные туловищные почки, от которых отходят мочеточники, открывающиеся в клоаку. В стенке клоаки имеется отверстие мочевого пузыря, в который стекает моча, попавшая в клоаку из мочеточников. В туловищных почках не происходит обратного всасывания воды. После наполнения мочевого пузыря и сокращения мышц его стенок, концентрированная моча выводится в клоаку и выбрасывается наружу. Часть продуктов обмена и большое количество влаги выделяется через кожу. Эти особенности не позволили земноводным полностью перейти к наземному образу жизни. У личинок на ранних стадиях развития функционирует т. н. головная почка, или предпочка. Также у всех амфибий лопастная печень, желчный пузырь, поджелудочная железа.
Половая система. Все земноводные раздельнополые. У большинства земноводных оплодотворение наружное (в воде). В период размножения наполненные зрелыми яйцеклетками парные яичники заполняют у самок почти всю брюшную полость. Созревшие икринки выпадают в брюшную полость тела, попадают в воронку яйцевода и, пройдя по нему, через клоаку выводятся наружу. Самцы имеют парные семенники. Отходящие от них семявыводящие канальца попадают в мочеточники, одновременно служащие самцам семяпроводами. Они также открываются в клоаку. Половые клетки по трубчатым протокам попадают в клоаку, а оттуда выбрасываются наружу.
0Головной мозг взрослой лягушки помещается в черепной коробке таким образом, что его наиболее широкая часть (зрительные доли) располагается между двумя ушными капсулами, или переднеушными костями. Обычные 5 отделов головного мозга - продолговатый мозг, мозжечок, средний мозг, промежуточный и передний - хорошо заметны только при рассмотрении со спинной стороны. Продолговатый мозг (medulla oblongata = myelencephalon) представляет собой непосредственное продолжение спинного. Его спинная стенка утончена, и перед ней в форме небольшого поперечного валика лежит мозжечок (cerebellum = metencephalon). Средний мозг (mesencephalon) несет на своей спинной поверхности округлые зрительные доли (lobi optici). С нижней стороны средний мозг образует мозговые ножки (реdunсuli cerebri), прикрытые нижним мозговым придатком и воронкой и ограниченные спереди зрительным трактом. Спинная крыша промежуточного мозга (diencephaIon = thalamencephalon) несет надмозговые придатки, а его нижняя поверхность -воронку.

Рис. 1. Головной мозг (а - сверху, б - сбоку, в - снизу). Римскими цифрами обозначен порядок головных нервов:
1 - затылочный полюс полушарий, 2- несимметричный ганглий, 3 - зрительный тракт, 4 - обонятельный нерв, 5 - обонятельные доли, 6- краевая ямка, 7 - стреловидная щель, 8-полушарие переднего мозга, 9 - передний надмозговой придаток, 10 - непарная спайка, 11 - задний надмозговой придаток (эпифиз), 12- промежуточный мозг, 13 - зрительная доля, 14 - мозжечок, 15 - слуховой гребень, 16 - промежуточная бороздка, 17- конечная пластинка, 18 - зрительная хиазма, 19 - воронка, 20 - нижний мозговой придаток (гипофиз), 21 - отхождение группы блуждающего нерва, 22 - нижняя часть полушарий, 23 - пучковая возвышенность, 24 - возвышенность между ножками, 25 - второй спинномозговой нерв.
К последней примыкают зрительная хиазма и нижний мозговой придаток. Границей промежуточного мозга спереди является концевая пластинка (lamina terminalis). Передний мозг (telencephalon) состоит из двух полушарий (hаеmisрhеriа), разделенных стреловидной щелью (fissurasagittalis) и сросшихся по средней линии обонятельных долей (lobi оlfасtorii). Последние внешне отделены от полушарий краевой ямкой (fovea limbic а). Для завершения первоначальной ориентировки полезно рассмотреть. От обонятельных долей переднего мозга отходят нервы 1 пары. Нервы II пары отходят от боковых стенок промежуточного мозга. Нервы III пары отходят от мозговых ножек среднего мозга. На границе между мозжечком и средним мозгом начинаются нервы IV пары. Все прочие головные нервы (V-X пары) отходят от продолговатого мозга.
Граница продолговатого мозга со спинным может быть приблизительно намечена только с брюшной стороны по отхождению II спинномозгового нерва. Передняя граница продолговатого мозга, напротив, хорошо отмечена со спинной стороны мозжечком, тогда как с брюшной стороны продолговатый мозг незаметно переходит в подмозжечковую область (rеgiо subcerebellaris). На нижней стороне хорошо видно продолжение срединной брюшной щели спинного мозга. Центральный канал спинного мозга расширяется в продолговатом мозгу в IV желудочек (ventriculus quartus). Последний прикрыт сверху тоненькой эпителиальной пластинкой (vellum mеdullаrе), лишенной нервных элементов. Эта пластинка образует массу складок, в которые заходит первичная мягкая мозговая оболочка, богатая кровеносными сосудами, -образуется заднее сосудистое сплетение (tela chorioi dea ventriculi quarti=plexus chorioideus posterior). При препаровке упомянутая эпителиальная пластинка часто удаляется вместе с мозговой оболочкой, и тогда через образовавшуюся «ромбовидную ямку» (fossa rhomboidalis) становится видимым дно IV желудочка, вдоль которого идет центральная бороздка (sulcus centralis).

Рис. 2. Нижний мозговой придаток:
1-третий головной нерв, 2 - передняя доля придатка, 3 - задняя доля придатка.

Рис. 3. Срединный продольный срез через головной мозг молодой лягушки (схема):
1 - конечная пластинка, 2 - передний надмозговой придаток, 3 - поводковая спайка, 4 - задний надмозговой придаток, 5 -- крыша промежуточного мозга, 6 - задняя спайка, 7 - непарный вырост, 8 - желудочек среднего мозга, 9 -заднее четыреххолмие, 10 - мозжечок, 11 -заднее сосудистое сплетение, 12- четвертый мозговой желудочек, 13 - обонятельная доля, 14- остаток невропора, 15 - конечная пласгинка, 16 - переднее сосудистое сплетение, 17 - Монроева щель, 18 - третий мозговой желудочек, 19 - воронка, 20 - передняя доля нижнего мозгового придатка, 21 - задняя доля нижнего мозгового придатка.
Бока этой ямки, стоящие в непосредственном отношении к VIII головному нерву, называют слуховым гребнем (eminentia acustica). От боковых стенок продолговатого мозга берут начало VII, IX и Х нервы.

Рис. 4. Схема основных пучков и путей головного мозга. Римскими цифрами обозначен порядок головных нервов:
1 - луковично-корковой, 2 - корково-спайковый, 3 - обратно изогнутый, 4 - корково-бугровый, 5 - зрительный, 6 - крыше-луковичный, 7 - мозжечково-спинномозговой, 8 - обонятельно-спайковый, 9 - обонятельный, 10 - мозжечково-промежуточный, 11 -луковично-спинной.
Нерв VI пары начинается от нижней поверхности продолговатого мозга. В заднюю часть продолговатого мозга продолжаются те образования, которые имеются в спинном мозгу, но к ним добавляется новообразование-уже упомянутый слуховой гребень. В массе серого вещества сперва уменьшаются, а потом и совсем исчезают брюшные рога. Такая же участь постигает и спинные рога, но от них сохраняется группа клеток, именуемая ядром спинного рога (nucleus cornus dorsalis). От этого ядра отходят к брюшной комиссуре внутренние дуговидные волокна (fibrae arcuatae intenae). Центральное серое вещество сперва диференцируется на таковое дна и крыши. Первое идет неизменно вдоль всего продолговатого мозга, а второе под влиянием раскрывающейся ромбовидной ямки постепенно отходит к бокам и исчезает. Через серое вещество крыши проходит изолированный тракт (tractus solitarius), достигающий наибольшей мощности в районе V нерва. Начиная с района II спинного нерва до района отхождения V головного нерва идет группа больших нервных клеток - центральное ядро (nucleus centralis). Что касается до белого вещества, то спинные пучки уплощаются еще в передней;части спинного мозга. Затем они постепенно отходят в стороны и в районе V нерва полностью исчезают. Срединный длинный пучок проходит через весь продолговатый мозг в средний. Через брюшную часть белого вещества проходит ряд трактов: мозжечково-спинной (tractus spino cerebellaris=cerebelIo-spinaIis), перешейково-спинной (t. isthmо - spinalis), корково-спинной (t. tecto-spinalis) и луковично-спинной (t. spinоthalamiсus = thalamo-spinalis). В районе VI нерва внутри белого вещества находится группа мелких клеток, стоящая в связи с серым веществом, -олива (oliva superior).


Рис. 5. Схематизированные поперечные срезы через продолговатый и спинной мозг: а - через спинной мозг тотчас позади отхождения II спинномозгового нерва, б - через задний конец продолговатого, тотчас перед отхождением II спинномозгового нерва, в - через продолговатый в районе группы блуждающего нерва, г - через продолговатый в районе отхождения VII и VIII нервов, д - не вполне симметричный через продолговатый в районе отхождения V нерва. Римскими цифрами обозначен порядок головных нервов:
1 - спинной пучок, 2 - боковой пучок, 3 - длинный срединный пучок, 4 - изолированный тракт, 5 - ядро спинного рога, 6 - спиннобоковые клетки, 7 - центральное ядро, 8 - срединнобрюшные клетки, 9 - подэпителиальная зона, 10- серое вещество слухового поля, 11- волокнистая масса слухового поля, 12 - периферия серого вещества, 13 - центральное серое вещество, 14 - двигательное ядро X нерва, 15 - наружные дуговидные волокна, 16 - мозжечково-спинномозговой тракт, 17 - двигательное ядро, 18 - брюшное слуховое ядро, 19 - спинное слуховое ядро, 20 - перерезанные волокна слухового поля, 21 - олива, 22 - ответвление слухового поля, 23 - двигательный корешок V нерва, 24 - двигательное ядро V нерва, 25 - зернистый слой мозжечка, 26 - чувствительный корешок V нерва.
Опыты показали, что лягушки, у которых были удалены все части головного мозга, лежащие впереди продолговатого, оставались в живых до 4 месяцев. При покойном сидении они держали конечности не так хорошо, как нормальные. Голова образовывала с телом тупой угол. У оперированных таким образом экземпляров наблюдалось непреодолимое стремление к передвижению. Изредка в ответ на внешнее раздражение наблюдались прыжки, сами же по себе животные только ползали или шагали. За добычей не охотились. Если надрезать и продолговатый мозг, то расстройства становятся тем значительнее, чем ближе к спинному мозгу сделан надрез. Если разрез приближается к началу плечевого сплетения, то двигательная способность передних конечностей все более нарушается, и животное уже полностью лежит на брюхе. Такое животное отвечает на раздражение в районе клоаки сильным ударом задних конечностей, но передние при этом лишь слабо дрожат.
Мозжечок (cerebellum = metencephalon) лягушки представляет собой небольшую, стоящую почти перпендикулярно к поверхности мозга поперечную пластинку, которая опирается на подмозжечковую область (regio subcerebellaris), в свою очередь являющуюся продолжением слухового гребня каждой стороны. Спереди мозжечок отграничен от среднего мозга поперечной складкой (fissura cerebri transversa). Дно этой складки называется передним мозговым парусом (velum mеdullare anterius). Задняя поверхность мозжечка обращена в четвертый желудочек. Подмозжечковая область идет вперед до возвышенности между ножками (eminentia interpeduncularis), а далее кпереди располагается «перешеек» («isthmus»). Общее расположение различных образований в подмозжечковой области и перешейке отвечает таковому продолговатого мозга, тогда как сам мозжечок имеет специфическую структуру. В мозжечке можно различить 6 слоев, следующих, друг за другом: эпителиальный, субэпителиальный зернистый, нервных волокон, передний зернистый, клеток Пуркинье и молекулярный. Назад от мозжечка идет уже упомянутый мозжечково-спинномозговой тракт (tractus сеrebello-spi nalis = spi nocerebellaris), а вперед - мозжечково-промежуточный (tractus cerebello-dience-рhаliсus). После удаления мозжечка положение тела и прыжки подопытных лягушек почти не отличаются от нормальных. Плавание протекает совершенно нормально, но если животное прыгает, то его прыжок оказывается либо слишком большим, либо слишком коротким для достижения цели. Взобравшаяся на край сосуда лягушка нередко остается в таком положении, при котором задняя часть ее тела более или менее перевешивается через край. Нормальная лягушка, напротив, успокаивается только тогда, когда она везде имеет под собой прочную опору.
Средний мозг (mesencephalon) сверху представлен двумя овальными зрительными долями, или «двухолмием» (lobi optici = corpora bigemina). Удаление зрительных долей лягушки вызывает слепоту. Нижнюю часть среднего мозга составляют большие парные массы продольных волокон - мозговые ножки (реdunсuli cerebri), отделенные от предшествующего раздела мозга только возвышенностью между ножками (еminеntia interpeduncularis). Ниже мозговых ножек располагается нижний мозговой придаток. Продолжение полости IV желудочка в средний мозг у высших позвоночных значительно суживается и получает название Сильвиева водопровода. У лягушки же это продолжение довольно обширно, и потому его лучше называть желудочком среднего мозга (ventriculus mesencephali). В нем нетрудно отличить две части -нижнюю и верхнюю. Последняя распадается на три выроста - один непарный (diverticulum imраr), направленный вперед, и два парных (diverticula lоbоrum), идущих в зрительные доли. Непарный вырост располагается над задней спайкой (comissura posterior).

Рис. 6. Схематизированные поперечные срезы через мозжечок; а - срез в районе выхода тройничного нерва, б - не вполне симметричный срез в районе перешейка:
7 - молекулярный слой, 2 - клетки Пуркинье, 3 - зернистый слой, 4 - эпителий, 5 - подэпителиальная зона, 6 - центральное серое вещество, 7 - остаток брюшного рога, 8 - мозжечок, 9 - волокна слухового поля, 10- мозжечково-спинномозговой тракт, 11- срединная бороздка, 12 - подмозжечковая область, 13 - срединный длинный пучок, 14 - задний корешок зрительного нерва, 15 - тракт поперечной спайки, 16 - ганглий перешейка, 17 - зрительная доля, 18 - блоковидный нерв, 19 - возвышенность между ножками.
В то время как в продолговатом мозгу столь много продольных волокон, что различение серого и белого вещества не вызывает затруднений, в переднем мозгу можно говорить только о богатом клетками внутреннем слое и бедном клетками наружном. Средний мозг занимает в этом отношении промежуточное положение, ибо в его нижней части (район мозговых ножек) еще можно назвать бедное клетками наружное вещество «белым». В крыше среднего мозга представителями белого вещества являются более толстый глубокий (stratum medulare profundum) и более тонкий поверхностный (str. mеdulare superficialis) мозговые слои. Вообще крыша среднего мозга достигает большой степени диференцировки, насчитывая до 9, а по некоторым авторам, до 15 слоев (изнутри наружу): эпителий, подэпителиальный зернистый, внутренний молекулярный, средний зернистый, наружный молекулярный, наружный зернистый, глубокий мозговой, промежуточный зернистый и поверхностный мозговой. Первые шесть из этих слоев отвечают понятию центрального серого вещества и составляют вместе богатый клетками внутренний слой. Поверхностный мозговой слой представляет собой продолжение подходящего спереди зрительного тракта (tractus opticus), тогда как глубокий есть продолжение элементов, пришедших из спинного мозга. В задней части верхнего раздела желудочка среднего мозгаимеется парное вздутие, которое некоторые называют задней частью четверохолмия («corpus quadrigeminum post. erius» = ganglion isthmi).

Рис. 7. Схематизированные поперечные срезы через средний мозг; а - через заднюю часть, б - в районе выхода глазодвигательного нерва, в - через переднюю часть:
1 - глубокий мозговой слой, 2 - передний корень зрительного нерва, 3 - задняя часть четверохолмия, 4 - крупные клетки наружного слоя, 5 - задний корень зрительного нерва, 6 - поперечная спайка, 7 - срединный длинный пучок, 8 - ядро глазодвигательного нерва, 9 - промежуточный зернистый слой, 10 - волокна внутреннего слоя, 11 - глазодвигательный нерв, 12- спайка крыши, 13 - эпителий, 14 - подэпителиальный зернистый слой, 15 - средний зернистый слой, 16 - наружный зернистый слой, 17 - глубокий мозговой слой, 18 - промежуточный зернистый слой, 19 - поверхностный мозговой слой, 20 - боковое глубокое ядро, 21- брюшная покрывающая спайка, 22 - спинная покрывающая спайка, 23 - непарный вырост, 24 - задняя спайка, 25 - крупноклеточное ядро, 26 - внесосковый ганглий, 27 - воронка.
Промежуточный мозг (diencephalon) виден только снизу, тогда как со спинной стороны он частично прикрыт соседними участками, а по бокам скрыт волокнами зрительных нервов. Его боковые части (partes subpalliales) переходят впереди в полушария, а между началами последних находится непарная концевая часть (parstеrminаlis), или концевая пластинка (lamina terminalis). На крыше промежуточного мозга находится ряд образований, называемых в совокупности надбугровйми (ерithа-lamus), тогда как образования, располагающиеся с нижней стороны промежуточного мозга, называются подбугровыми (hypothalamus). Соответственно утолщенные боковые стенки именуются зрительными буграми (thаlamiорtiсi). Задние две трети боковых стенок промежуточного мозга одеты волокнами зрительного тракта, расходящимися от перекреста (хиазмы), расположенного снизу. Главная масса этих волокон направляется к зрительным долям, где и распадается на спинной, промежуточный и брюшной корни (radix dorsalis, r. intermedia, r. ventralis). Часть боковых стенок промежуточного мозга, лежащая впереди волокон зрительного тракта, вздута в пучковую возвышенность (prominentia fascicularis). Надбугровыми образованиями являются два непарных надмозговых придатка. Передний надмозговой придаток (paraphysis = nodulus vasculosus) представляет собой небольшой узелок, богато снабженный кровеносными"сосудами. На продольном срезе видно, что основание его вдается в переднюю часть полости мозгового желудочка в виде сосудистого сплетения (plexus chorioideus). Задний надмозговой придаток (epiphysis) имеет вид тонкой нити (pedunculus corporis pinealis), оканчивающейся небольшим тельцем (corpus pineal е) на поверхности черепа. Между основаниями переднего и заднего надмозговых придатков проходит поперек мозга поводковая, или верхняя спайка (соmissura habenularis = c. superior), по бокам которой лежит по небольшому поводковому бугорку (ganglion habenulae). Переходя кподбугровым образованиям, надо отметить, что впереди зрительного перекреста располагается основная часть (pars basаlis) концевой пластинки, а сзади - доля воронки (lоbus infundibularis=pars postchiasmatica hypothalami). Последняя есть результат непарного выроста дна третьего желудочка - воронки (infundibulum). Позади доли воронки находится нижний мозговой придаток (hypophysis = glandula pituitaria), имеющий, однако, совсем другое происхождение (из крыши ротовой полости) и являющийся органом внутренней секреции. Нижний мозговой придаток состоит из двух частей: небольшой трехраздельной передней (раrs anterior) и более крупной задней (pars роstеriоr). На продольном разрезе видно, что впереди зрительного перекреста дно III мозгового желудочка дает зрительную впадину (recessus opticus). Перед этой впадиной поперек продольной оси мозга идут две передних спайки (comissura pallii anterior et comissura anterior). Что касается более тонкой структуры, то и в тех частях промежуточного мозга, которые не находятся в примитивном эпителиальном состоянии, можно различить три слоя: эпителий, богатый клетками внутренний слой (центральное серое вещество) и бедный клетками наружный слой. Часть последнего, пронизанная волокнами зрительного нерва, называется коленчатыми телами (corpora geniculata).

Рис. 8. Схематизированные срезы через промежуточный мозг; а - сзади зрительного перекреста, б - в районе зрительного перекреста, в - в переднем отделе:
1 - ножка заднего надмозгового придатка, 2 - верхняя ограничивающая зона, 3 - нижняя ограничивающая зона, 4 - коленчатое тело, 5 - основной пучок переднего дюзга, 6 - зрительный слой, 7 - срединный пучок переднего мозга, 8 - надвороночный перекрест, 9 - верхняя часть серого слоя, 10 - глубокий мозговой слой, 11 - средняя часть серого слоя, 12 - нижняя часть серого слоя, 13- доля воронки, 14 - поводковая спайка, 15 - поверхностное переднее ядро, 16- нижний слой зрительного перекреста, 17-верхний слой зрительного перекреста, 18 - поводковый узел, 19 - передний надмозговой придаток, 20 - узел перегородки, 21 - боковой желудочек, 22 - третий желудочек, 23 - передняя спайка, 24 - зрительный нерв, 25 - спинное образование мантии, 26 - боковое образование мантии, 27 - срединное образование мантии, 28 - верхняя часть передней спайки, 29 - пучковая возвышенность, 30 - нижняя часть передней спайки, 31 - зрительная впадина, 32 - затылочный полюс, 33 - дуговая щель.

Рис. 9. Схематизированные поперечные срезы через передний мозг; а - в районе Монроевых щелей, б - в задней части полушарий, в - через середину полушарий, г - в средней части обонятельных долей:
1- передний надмозговой придаток, 2 - перегородка, 3 - боковой желудочек, 4 - передняя спайка мантии, 5 - Монроева щель, 6 - зрительный нерв, 7 - основной пучок переднего мозга, 8 - срединный пучок переднего мозга, 9 - дуговая щель, 10 - боковое образование мантии, 11 - срединное образование мантии, 12 - ограничивающая зона, 13 - основной узел, 14 - конечная часть, 15 - зрительная впадина, 16 - промежуточная бороздка, 17 - боковая ограничивающая бороздка, 18 - узел перегородки, 19 - стреловидная щель, 20 - граница мантии и подмантии, 21 - задний корень обонятельного нерва, 22 - поверхностный волокнистый слой, 23 - продолжение бокового желудочка, 24 - зернистый слой, 25 - молекулярный слой, 26 -крупноклеточный слой, 27 - слой клубочков,
Передний, или конечный, мозг (telencephalon) состоит из двух полушарий и обонятельных долей. Каждое полушарие (hаеmisphaerium) представляется при взгляде сверху овальным телом, несколько суживающимся кпереди. Правое и левое полушария, как уже указывалось, спереди ограничены краевой ямкой (fossa limbiса), а друг от друга отделены стреловидной щелью (fissura sagittalis). При рассматривании сбоку на полушарии можно различить две части: верхнюю выпуклую - мантию (pars superior - p. pallialis = pallium) и нижнюю, или подмантийную (pars inferior = p. subpal lialis = subpallium). Последняя является прямым продолжением нижней и боковых стенок промежуточного мозга. От мантии подмантийную часть отделяет неглубокая боковая ограничивающая бороздка (sulcus limitanslateralis). С внутренней стороны каждого полушария аналогичной границей является дуговая щель (fissura arcuata - sерti). Часть подмантии, расположенная ниже дуговой щели, носит название перегородки (septum).
В задней части срединной поверхности полушарий видна еще одна очень слабая продольная бороздка, идущая ниже дуговой. Отграниченное ими пространство отвечает положению узла перегородки (ganglium mediate septi). Мантия своим задним концом-затылочным полюсом (polus occipitalis) несколько налегает на промежуточный мозг. Внутри каждого полушария находится боковой желудочек (ventriculus haemisphaerii = v. lateralis), сообающийся с II мозговым желудочком посредством межжелудочного отверстия, или Монроевой щели (foramen interventricularе - f. Моnrоi). Начиная от этого отверстия, вдоль внутренней поверхности срединной стенки полушария идет промежуточная бороздка (sulcus intermedius). Спереди она загибается кверху и получает наименование предмантийной (sulcus ргаеpallialis). Разграниченные промежуточной бороздкой части срединной стенки полушария вдаются внутрь бокового желудочка под названием срединного возвышения мантии (еminеntiapallialis medialis) и возвышения перегородки (eminentia septalis), лежащего ниже упомянутой бороздки. Продолжение последней, лежащее впереди предмантийной бороздки, называется заобонятельной возвышенностью (eminentia postolfасtоriа). На поверхности данной возвышенности имеется слабая предкраевая бороздка (sulcus praelimbicus). Наружная (боковая) стенка каждого полушария тоньше, чем срединная, и почти лишена особого рельефа. В стенках полушарий лягушки еще нет той диференцировки, которая наблюдается у вышестоящих форм. В стенках полушарий можно различить два слоя: богатый клетками внутренний и бедный клетками наружный. Отношение этих слоев друг к другу может меняться, и во внутреннем слое местами формируются особые «образования» (fоrmatiо), несколько отличные по своему клеточному строению. Так, например, в мантии различают срединное образование (formatio pallialis medialis), спинное (f. p. dorsaIis) и боковое (f. p. lateralis). В первом из них обнаружено три клеточных типа: большие пирамидальные клетки, клетки с многочисленными расходящимися во все стороны отростками и тангенциальные клетки. Подобные элементы встречаются и в спинном образовании мантии, но лежат в нем много теснее. Наконец, боковое образование есть небольшой слой, клетки которого сидят тесно и правильно.

Рис. 10. Срединная стенка левого полушария, видимая благодаря удалению боковой стенки. На. срезе обонятельной доли нанесены клеточные слои:
1 - промежуточная бороздка, 2 - предмантийная бороздка, 3 - заобонятельная возвышенность, 4 - предкраевая бороздка, 5 - обонятельный нерв, б - поверхностный волокнистый слой, 7 - слой клубочков, 5 - крупноклеточный слой, 9 - молекулярный слой, 10 - зернистый слой, 11 - срединная возвышенность мантии, 12 - затылочный полюс, 13 - Монроева щель, 14 - возвышенность перегородки.
Обонятельные доли (lobi olfactoril) примыкают непосредственно к полушариям. Срединные части обеих долей столь тесно срастаются друг с другом, что их самостоятельность может быть установлена либо на поперечном срезе по раздельным продолжениям в них полостей боковых желудочков, либо по слабым продольным бороздкам на их верхней и нижней поверхности - спинной срединной (sujcus medianus dorsalis) и брюшной срединной (sulcus medianus ventralis). Впереди обонятельные доли переходят в обонятельные нервы. Однако каждый из них отходит еще дополнительным, более слабым, задним или боковым корнем от дополнительной обонятельной луковицы (bulbusоlfа-ctorius accessorius). Расположение клеточных слоев обонятельных долей отличается от такового полушарий некоторыми деталями. Утолщенный в ряде мест внутренний слой получает название зернистого (stratum granulosum). Вторжение в обонятельные доли нитей обонятельного нерва приводит к образованию новых слоев (снизу вверх): поверхностного волокнистого (stratumfib-rillare superficiale), слоя клубочков (str. glоmerulоsum) и крупноклеточного (st. magnосellulare).
Главными проводящими путями переднего мозга будут: обратноизогнутый (fasciculus retroflexus), корково-спайковый (f. corti cohabenularis), корково-бугровый (f. соrticothalamicus), обонятельный (tractus olfactorius) обонятельно-спайковый (tr. olfactohabenularis) и луковично-корковый (tr. bulbосоrtiсalis).
Удаление полушарий без повреждения промежуточного мозга не дает заметных расстройств, за исключением утраты обоняния. Лягушки самопроизвольно прыгают с места на место, нормально плавают, а с наступлением холодов прячутся под камни или в землю.
Избегают врага и ловят добычу. Отмечают, что оперированные таким образом лягушки регулярно квакают при поглаживании по спине. Напротив, при одновременном повреждении промежуточного мозга лягушка утрачивает всякую самостоятельность. Не побуждаемая внешним раздражением к передвижению, такая лягушка сидит совершенно неподвижно, пока не высохнет как мумия. Все движения ее сходны с движениями нормальной лягушки, но представляются совершенно машинальными и единообразными. Если погружать такую лягушку в воду, то она не оказывает никакого сопротивления. При полном удалении головного мозга рефлекторная возбудимость повышается (снятие мозгового торможения).
Центральная нервная система лягушки окружена тремя мозговыми оболочками (meningеs). Непосредственно к костям и хрящам прилегает изнутри тонкая надкостница, или надхрящница (реriosteum = «endorachis»). Под ней помещается наполненное лимфой околооболочечное пространство (cavum periduralе), за которым следует крепкая фиброзная твердая оболочка (dura mater). Подфиброзное пространство (cavum subduralе) отделяет последнюю от примитивной мягкой оболочки (рia mater primitivа). При вскрытии полостей, где помещается центральная нервная система, надкостница обычно разрывается, и наблюдателю открывается упомянутое околооболочечное пространство. Мягкая оболочка, несущая кровеносные сосуды, в некоторых местах (брюшная щель, район концевой нити) расщепляется на два листка.

Рис. 11. Места выхода головных нервов. Вид на правую половину черепа изнутри. Римскими цифрами обозначен порядок нервов:
1 - переднеушная кость, 2 - общий- переднеушной узел, 3 - эндолимфатическое отверстие, 4 - переднеушно-затылочный синхондроз, 5 - боковая затылочная кость, 6 - группа блуждающего нерва, 7 - обонятельное отверстие, 8 - обонятельная кость, 9 -симпатический нерв.
Используемая литература: П. В. Терентьев
Лягушка: Учебное пособие/ П.В. Терентьев;
под ред. М. А. Воронцова, А. И. Прояева.- М. 1950 г.
Скачать реферат: У вас нет доступа к скачиванию файлов с нашего сервера.
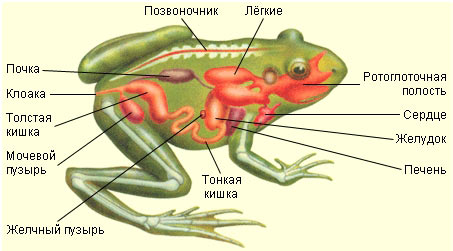
137. Рассмотрите рисунки. Напишите названия частей тела лягушки. Какие органы расположены на ее голове? Напишите их названия.
138. Изучите таблицу "Класс Земноводные. Строение лягушки". Рассмотрите рисунок. Напишите названия внутренних органов лягушки, обозначенных цифрами.
3. желудок
4. поджелудочная железа
5. семенник
8. толстая кишка
9. тонкая кишка
10. желчный пузырь
11. печень
139. Опишите строение головного мозга земноводных.
Головной мозг у земноводных имеет более прогрессивные черты, более крупные размеры переднего мозга, полное разделение полушарий. Средний мозг относительно небольшой. Мозжечок мал, так как у амфибий однообразные движения. От головного мозга отходят 10 черепно-мозговых нервов.
Отделы головного мозга: передний, средний, мозжечок, продолговатый, промежуточный.
140. Изучите таблицу "Класс Земноводные. Строение лягушки". Рассмотрите рисунок. Подпишите названия частей скелета лягушки, обозначенных цифрами.

2. лопатка
4. предплечье
9. уростиль
10. позвоночник
141. Рассмотрите рисунок. Напишите названия пищеварительной системы лягушки, обозначенных цифрами. Как осуществляется процесс пишеварения у лягушки?

1. ротовое отверстие
2. пищевод
3. желудок
4. кишечник
Все земноводные питаются итолько подвижной пищей. На дне ротовой полости находится язык. При ловле добычи он выкидывается изо рта, к нему прикрепляется добыча. На верхней челюсти имеются зубы, служащие только для удерживания добычи. При глотании проталкиванию пищи в пищевод помогают глазные яблоки.
В ротоглотку открываются протоки слюнных желез. Из ротоглотки пища по пищеводу попадает в желудок, оттуда в 12-перстную кишку. Сюда открываются протоки печени и поджелудочной железы. Переваривание происходит в желудке и 12-перстной кишке. Тонкий кишечник переходит в прямую, которая образует расширение - клоаку.
142. Зарисуйте схему строения сердца лягушки. Какую кровь называют артериальной, а какую - венозной?
Артериальная кровь идет от легких, богата кислородом. Венозная - к легким.

143. Опишите процесс размножения и развития лягушки. Укажите сходство и различие в размножении земноводных и рыб.
Размножаются земноводные в небольших хорошо прогреваемых участках водоемов. Органы размножения у самцов - семенники, у самок - яичники. Оплодотворение наружное.
Развитие лягушки: яйцо - головастик в момент выклева - развитие плавниковых складок и наружных жабр - стадия максимального развития наружных жабр - стадия исчезновения наружных жабр - стадия появления задних конечностей - стадия расчленения и подвижности задних конечностей - стадия освобождения передних конечностей, метаморфоза ротового аппарата и начала резорбции хвоста - стадия выхода на сушу.
144. Заполните таблицу.
Строение и значение органов чувств лягушки.
Органы чувств лягушки Особенности строения Значение Глаза Есть верхние (кожистые) и нижние (прозрачные) подвижные веки, мигательная перепонка. Есть железа, секрет которой смачивает роговицу и предохраняет ее от высыхания. Роговица выпуклая. Хрусталик имеет форму двояковыпукло линзы. У многих развито цветное зрение Зрение Орган слуха Внутренне ухо, среднее ухо. Наружно слуховое отверстие закрывает барабанная перепонка, содиненная со слуховой косточкой - стремячом Слух Орган равновесия Внутренне ухо Ориентировка Орган обоняния Парные обонятельные мешки. Их стенки выстланы обонятельным эпителием. Открываются наружу ноздрями, а в ротоглотку - хаонами Восприятие запахов Орган осязания Кожа Восприятие раздражения Орган боковой линии Боковая линия у личинок Позволяет ощущать ток воды
СТРОЕНИЕ ЗЕМНОВОДНЫХ
Кожные покровы . Кожа у всех земноводных голая, лишенная чешуи. Эпидермис богат многоклеточными железами (рис. 68). Значение кожных желез многообразно. Они обеспечивают наличие на поверхности кожи жидкостной пленки, без которой при кожном дыхании невозможен газообмен. Эта пленка в некоторой мере предохраняет тело от иссушения. Слизь помогает коагуляции инородных частиц на поверхности тела. Выделения некоторых кожных желез обладают бактерицидными свойствами и защищают тело от проникновения через кожу патогенных микробов. Ядовитые кожные железы в значительной степени защищают амфибий от хищников. Часто ядовитые формы имеют яркую окраску,
Рис. 68. Разрез через кожу саламандры:
1 - выступающий наружу секрет железы; 2 - пигментный слой;3 - слизистые кожные железы; 4 - ядовитая кожная железа; 5 – перерезанные кровеносные сосуды, 6 - эпидермис; 7 - волокнистый слой кожи
предупреждающую хищников об опасности при схватывании добычи. Установлено, что у некоторых бесхвостых амфибий верхний слой эпидермиса ороговевает. Наиболее сильно это развито у жаб, роговой слой у которых на спине составляет примерно 60% всей поверхности эпидермиса. У большинства амфибий слабое ороговение эпидермиса не препятствует проникновению через кожу воды, и, находясь в водоеме, амфибии постоянно "пьют воду кожей".
Скелет земноводных по сравнению с рыбами имеет ряд изменений. В осевом скелете позвоночник земноводных в связи с полуназемным образом жизни более расчленен. Он включает шейный, туловищный, крестцовый и хвостовой отделы (рис. 69). Шейный отдел представлен одним позвонком, его тело невелико и несет две сочленовные ямки, при помощи которых позвонок сочленяется с черепом. Число туловищных позвонков различно. Наименьшее число их у бесхвостых (обычно 7), наибольшее - у безногих (более 100). Единственный крестцовый позвонок (у безногих отсутствующий) несет длинные поперечные отростки, к которым причленяются подвздошные кости таза. Хвостовой отдел наиболее типично выражен у хвостатых, у безногих он очень мал, а у бесхвостых представлен косточкой - уростилем : во время эмбрионального развития он закладывается в виде ряда отдельных позвонков, впоследствии срастающихся.
Форма позвонков у разных представителей в пределах класса земноводных чрезвычайно вариабельна. У низших амфибий (безногие, низшие хвостатые) она бывает амфицелъной в этом случае между позвонками пожизненно сохраняется хорда. У бесхвостых позвонки процелъные , т.е. вогнутые спереди и выгнутые сзади; у
высших хвостатых - опистоцелъные , т.е. выгнутые спереди и вогнутые сзади. Вариантов отступления от данной характеристики много, к примеру у крайне примитивной новозеландской лягушки леопелъмы (Leopelma) позвонки амфицельные. Настоящие ребра у бесхвостых не развиваются, очень короткие есть у безногих амфибий; у хвостатых развиваются короткие "верхние" ребра.
Мозговой череп. Мозговая коробка в значительной своей части пожизненно остается хрящевой (рис. 70). Это обусловливается слабым развитием хондральных и накладных окостенений. В первичном мозговом черепе развиваются следующие хондральные кости. В затылочной области только две боковые затылочные костищ места, соответствующие основной и верхней затылочным костям рыб, остаются хрящевыми. В области слуховой капсулы формируется одна небольшая ушная кость , большая же часть капсулы остается хрящевой. В передней части глазницы у бесхвостых развивается одна клинообонятельная кость , у хвостатых эта кость парная. Обонятельная капсула хрящевая.
Покровных костей также немного. Крышу черепа составляют теменные и лобные кости, которые у бесхвостых срастаются в лобно-теменные кости . Впереди от них находятся носовые кости, у безногих они срастаются с предчелюстными костями. По бокам задней части черепа расположены чешуйчатые кости , особенно сильно развитые у безногих. Дно черепа выстилает большой парасфеноид , а впереди от него лежат парные сошниковые кости.
В формировании дна черепа принимают участие и кости висцерального скелета - нёбные и крыловидные . Первые прилегают к

Рис. 69. Позвоночный столб лягушки вместе с тазовым поясом (со спинной стороны):
I - шейный отдел (из одного позвонка), II - туловищный отдел; III - крестец, IV - уростиль, 1 -остистый отросток 3-го туловищного позвонка, 2 - поперечный отросток того же позвонка, 3 - сочленовные поверхности на первом шейном позвонке

Рис. 70. Череп лягушки:
I - вид сверху; II - вид снизу (накладные кости с одной стороны удалены). Обозначение костей: 1 боковая затылочная, 2 - лобно-теменная; 3 - носовая; 4 - межчелюстная, 5 - переднеушная; 6 - чешуйчатая; 7 - парасфеноид (левая его половина); 8 - сошник; 9 - нёбная; 10 - крыловидная; 11 - клинообонятельная; 12 - верхнечелюстная; 13 - квадратночелюст-ная; 14 - отверстие для выхода зрительного нерва, 15 - отверстие для тройничного нерва
сошникам, вторые - к чешуйчатым костям. Они развиваются на нижней поверхности нёбно-квадратного хряща. Функции верхних челюстей выполняют, как у костных рыб, предчелюстные (или межчелюстные ) и верхнечелюстные кости.
Нижняя челюсть представлена меккелевым хрящом, который прикрыт снаружи зубной и угловой костями.
Череп амфибий аутостилический , т.е. нёбно-квадратный хрящ непосредственно прирастает к мозговому черепу. В связи с ауто-стилией черепа подъязычная дуга не принимает участия в прикреплении челюстного аппарата к черепу. Верхний элемент этой дуги -подвесок (гиомандибуляре) - превращен в маленькую кость - стремя , которая проксимальным концом упирается в слуховую капсулу, а наружним (дистальным) - в барабанную перепонку. В связи с формированием полости среднего уха эта кость находится внутри указанной полости и выполняет роль слуховой косточки. Таким образом, гиомандибуляре (подвесок) выходит из системы четвертой (подъязычной) висцеральной дуги (рис. 70).
Нижние элементы подъязычной дуги и жаберных дуг видоизменяются в подъязычную пластинку и ее рожки. Эта пластинка расположена между ветвями нижней челюсти. Передние ее рожки,

Рис. 71. Плечевой пояс лягушки спереди:
1 - грудина; 2 - хрящевые передняя и задняя части грудины; 3 - коракоид; 4 - ключица лежит на прокоракоиде;5 - предгрудина; 6 - лопатка; 7 - сочленовная впадина на лопатке для плеча (хрящ покрыт точками)
загибаясь вверх и охватывая с боков кишечную трубку, прикреплены к слуховым капсулам. Изменения в висцеральном скелете сопровождаются утратой жаберных крышек.
Таким образом, череп амфибий отличается от черепа большинства костных рыб: 1) слабым развитием хондральных и кожных окостенений; 2) аутостилией; 3) видоизменением подъязычной и жаберных дуг, превращенных частью в слуховой, частью в подъязычный аппарат; 4) редукцией жаберной крышки.
Пояса конечностей . Плечевой пояс имеет вид дуги, обращенной вершиной к брюшной поверхности животного (рис. 71). Каждая половина дуги (левая и правая) состоит из следующих основных элементов. Верхняя (спинная) часть представлена лопаткой с широким надлопаточным хрящом . Нижняя (брюшная) часть включает коракоид и лежащий впереди него прокоракоид . У бесхвостых между предгрудинои и лопаткой расположена тонкая палочковидная ключица . Перечисленные элементы пояса сходятся в точке прикрепления плечевой кости и формируют сочленовную ямку. Впереди от места соединения левого и правого коракоидов находится предгрудина , а сзади - грудина . Обе эти кости заканчиваются хрящами. Плечевой пояс в отличие от костных рыб лежит свободно в толще мускулатуры и не связан с черепом. В связи с отсутствием или неполным развитием ребер грудной клетки у амфибий нет.
Тазовый пояс (рис. 72) образован тремя парными элементами, сходящимися в области вертлужной впадины, .которую они и образуют. Длинные подвздошные кости своими проксимальными (передними) концами прикреплены к поперечным отросткам единственного крестцового позвонка. Направленный вперед и вниз лобковый элемент пояса у лягушек остается хрящевым. Сзади него расположена седалищная кость. Такое расположение элементов тазового пояса характерно для всех наземных позвоночных.
Скелет свободных конечностей типичен для наземных позвоночных и существенно отличен от скелета конечностей рыб. В то время как конечности рыб представляют в схеме простые

Рис. 72. Скелет лягушки:
I - целый скелет; II - позвонок сверху; III - позвонок спереди: 1 - шейный позвонок; 2 - крестцовый позвонок; 3 - уростиль; 4 - грудина; 5 – хрящевая задняя часть грудины; 6 - предгрудина; 7 - коракоид; 8 - прокоракоид; 9 - лопатка; 10 - надлопаточный хрящ; 11 - подвздошная кость; 12 - седалищная кость; 13-лобковый хрящ; 14-плечевая кость; 15 - предплечье (лучевая + локтевая кости); 16 - запястье; 17 - пясть; 18 - зачаточный I палец; 19 - II палец; 20 - V палец; 21 - бедро; 22 - голень (большая и малая берцовые кости); 23 - предплюсна; 24 - плюсна; 25 - рудимент добавочного пальца;26-1 палец; 27 - тело позвонка; 28 - спинномозговой канал; 29 – сочленовная площадка; 30 - остистый отросток; 31 - поперечный отросток
одночленные рычаги, перемещающиеся только относительно корпуса тела и не несущие мускулатуры, конечности наземных позвоночных представляют собой многочленные рычаги с достаточно мощными мышцами. В этом случае не только вся конечность перемещается относительно тела, но и отдельные элементы конечности перемещаются друг относительно друга.
В схеме пятипалая конечность состоит из трех основных отделов (рис. 72).
I - плечо в передней конечности, бедро - в задней; этот отдел всегда состоит из одной кости, которая проксимальным концом причленена к поясу;
II - предплечье в передней конечности, голень в задней. В типичном случае отдел состоит из двух параллельно расположенных
костей: предплечье - из локтевой и лучевой , голень - из большой берцовой и малой берцовой ,
III - кисть в передней конечности и стопа в задней; отдел состоит из трех подотделов:
1) запястье - в передней конечности, предплюсна - в задней; этот подотдел представлен в типичном случае 9-10 мелкими косточками, расположенными в три ряда;
2) пясть - в передней конечности, плюсна - в задней; в типичном случае подотдел состоит из 5 удлиненных костей, расположенных в один ряд, как бы веером, от запястья или предплюсны;
3) фаланги четырех-пяти пальцев представляют как бы продолжение пясти или плюсны и включают три-пять рядов косточек в каждом.
Скелет конечностей хвостатых амфибий почти полностью соответствует приведенной схеме. У лягушек наблюдаются некоторые отклонения. Основные из них следующие: оба элемента предплечья и голени срастаются в одну кость, срастается между собой и большая часть костей запястья и предплюсны, перед первым пальцем задней конечности имеется рудимент добавочного пальца. Эти особенности имеют вторичный характер и связаны, видимо, с приспособлением лягушек к передвижению прыжками.
Мышечная система существенно отличается от мышечной системы рыб двумя основными особенностями, связанными с передвижением животных при помощи пятипалых конечностей и в известной мере по твердому субстрату. Во-первых, развивается мощная и сложно организованная мускулатура на свободных конечностях. Во-вторых, в связи со сложными движениями мускулатура тела более дифференцирована, а характерная для рыб ее сегментация у амфибий нарушена. Метамерия мышечной системы выражена более четко у хвостатых и безногих. В свою очередь у бесхвостых ее можно увидеть только в немногих местах туловища у взрослых форм и в личиночном состоянии.
Органы пищеварения . Ротовая щель ведет в обширную ротоглоточную полость , которая, сужаясь, переходит в пищевод. В ротоглоточную полость открываются хоаны, евстахиевы отверстия (полости среднего уха) и гортанная щель . Сюда же открываются отсутствующие у рыб протоки слюнных желез . Их секрет служит для смачивания пищевого комка и не воздействует на пищу химически. На дне ротоглоточной области расположен настоящий язык , имеющий собственную мускулатуру. Форма языка разнообразна. У некоторых хвостатых он прикреплен неподвижно, у других имеет вид гриба, сидящего на тонкой ножке. У лягушек язык прикреплен одним концом ко дну рта, а свободная его часть в спокойном состоянии обращена внутрь. У всех амфибий язык
выделяет клейкое вещество и служит для ловли мелких животных. Немногие амфибии не имеют языка.
На межчелюстных и верхнечелюстных костях, на сошнике, а у некоторых и на нижней челюсти сидят зубы . Они имеют вид мелких однообразных конусов, вершины которых несколько загнуты назад. Некоторые виды, например жабы, зубов на костях челюстей не имеют. При глотании проталкиванию пищевого комка из ротоглоточной области в пищевод помогают глазные яблоки, которые отделены от этой полости только тонкой слизистой оболочкой и с помощью специальных мышц могут несколько втягиваться внутрь ротоглотки.
Короткий пищевод впадает в сравнительно слабо отграниченный желудок . Собственно кишечник относительно более длинный, чем у рыб. В петле переднего (тонкого ) отдела лежит поджелудочная железа . Крупная печень имеет желчный пузырь , ее проток впадает в переднюю часть тонкой кишки (в двенадцатиперстную кишку). В желчный проток впадают и протоки поджелудочной железы, которая самостоятельного сообщения с кишечником не имеет (рис. 73). Второй отдел кишечника - толстый - отграничен от тонкого отдела нечетко. Наоборот, третий, прямой отдел обособлен хорошо. Он заканчивается клоакой .

Рис. 73. Общее расположение внутренностей самки лягушки:
1 - пищевод; 2 - желудок; 3 - лопасти печени; 4 - поджелудочная железа; 5 - тонкая кишка; 6 - толстая (прямая) кишка; 7 - клоака (вскрытая); 8 - желудочек сердца;9 - левое предсердие; 10 - правое предсердие; 11 - сонная артерия (правая); 12 – левая дуга аорты; 13 - легочно-кожная дуга (правая); 14 - задняя (нижняя) полая вена; 15 - брюшная вена; 16 - легкие; 17 - левая почка (задний ее конец); 18 - правый яичник; 19 - левый яйцевод; 20 - его устье (воронка); 21 - мочевой пузырь; 22 - желчный пузырь; 23 - селезенка; 24 - передняя полая вена (правая)
Органы дыхания амфибий разнообразны. Во взрослом состоянии большинство видов дышат легкими и через кожу. Легкие представляют собой парные мешки с тонкими ячеистыми стенками. При относительно малой поверхности легких очень велико значение кожного дыхания . Отношение поверхности легких к поверхности кожи у амфибий равно 2:3 (в то время как у млекопитающих внутренняя поверхность легких в 50-100 раз больше поверхности кожи). У зеленой лягушки через кожу поступает 51% кислорода. Важна роль кожи в выделении из организма углекислоты: через кожу выделяется 86%, через легкие - 14%.
Кожное дыхание имеет большое функциональное значение не только в связи с несовершенством легких, но и как приспособление, обеспечивающее окисление крови при длительном нахождении животного в воде, например во время спячки или затаивания в водоеме при преследовании наземными хищниками. В этих случаях осуществляется только кожное дыхание, и правое предсердие (куда через полую вену вливается окисленная кровь кожной вены -см. ниже) становится артериальным, а левое - венозным.
У американских безлегочных саламандр и у дальневосточного тритона легкие полностью атрофированы, и газообмен у них целиком происходит через кожу и слизистую ротовой полости.
Способность кожи и легких поглощать кислород (О 2) и выделять углекислый газ (CO 2) зависит у амфибий от температуры среды. В таблице 5 представлены данные для зеленой лягушки.
Как видно, при температуре окружающей среды +5°С через кожу поступает в 1,5 раза больше кислорода, чем через легкие. При температуре +25°С, наоборот, через легкие поступает кислорода в 2 раза больше, чем через кожу. Иначе обстоит дело с выходом СO 2 . При повышении температуры от +5 до +25 °С значение кожи в выходе углекислого газа возрастает только в 3,3 раза, а легких -в 7 раз. При любых температурах роль кожи для удаления СО 2 заметно больше, чем легких.
Таблица 5
Зависимость потребления О 2 и выделения CO 2 от температуры среды у зеленой лягушки
(по Проссеру, 1977)
Личинки амфибий дышат при помощи ветвистых наружных жабр, которые у подавляющего большинства видов в последующем исчезают, а у протеев и сирен сохраняются пожизненно. Амфиумы во взрослом состоянии наряду с легкими имеют и внутренние жабры.
В связи с отсутствием грудной клетки механизм легочного дыхания весьма своеобразен. Роль насоса выполняет ротоглоточная полость, дно которой то опускается (воздух при открытых ноздрях засасывается), то поднимается (воздух при закрытых ноздрях проталкивается в легкие). Поэтому череп амфибий при небольшой высоте чрезвычайно широк: эффективность легочного дыхания тем больше, чем больше расстояние между ветвями нижней челюсти. К примеру, наибольшей ширины оно достигает у жаб, ороговевшая кожа которых имеет малую дыхательную нагрузку.
Кровеносная система . Сердце у всех амфибий трехкамерное, состоит из двух предсердий и одного желудочка (рис. 74). У низших форм (безногие и хвостатые) левое и правое предсердия разделены не полностью. У бесхвостых перегородка между предсердиями полная, но у всех земноводных оба предсердия сообщаются с желудочком одним общим отверстием. Кроме указанных основных отделов сердца имеется венозная пазуха . Она принимает венозную кровь и сообщается с правым предсердием. К сердцу примыкает артериальный конус , в него кровь изливается из желудочка. Артериальный конус имеет спиральный клапан , участвующий в распределении крови в выходящие из него три пары сосудов. Сердечный индекс (отношение массы сердца к массе тела в процентах) варьирует и зависит от двигательной активности животного. Так, у сравнительно мало перемещающихся травяной и зеленой лягушек он равен 0,35-0,55%, а у полностью наземной (кроме периода размножения) и активной зеленой жабы - 0,99%.
У высших - бесхвостых - амфибий от артериального конуса берут начало три пары артериальных дуг .
Первая пара (считая от головы к хвосту) несет кровь к голове, это - сонные артерии . Они гомологичны первой паре жаберных артерий рыб. Вторая пара, также отходящая от брюшной стороны артериального конуса, гомологична второй паре жаберных сосудов рыб и носит название системных дуг аорты . От нее отходят подключичные артерии , несущие кровь к плечевому поясу и передним конечностям. Правая и левая системные дуги, описав полукруг, соединяются вместе и образуют спинную аорту , расположенную под позвоночником и дающую начало артериям, идущим к внутренним органам. Последняя, третья, пара, гомологичная четвертой паре жаберных артерий рыб, отходит не от брюшной, а от спинной стороны артериального конуса. Она несет кровь в легкие и представляет собой легочные артерии . От каждой легочной
артерии отходит сосуд, несущий венозную кровь в кожу: это кожные артерии .
У хвостатых амфибий, имеющих легкие, схема расположения артериальных сосудов в основном такая же. Но, в отличие от бесхвостых, у них сохраняется пара артерий, соответствующая третьей паре жаберных сосудов, и, таким образом, общее число парных артериальных стволов у них равно четырем, а не трем, как у бесхвостых. Кроме того, легочные артерии сохраняют связь с системными дугами аорты через посредство так называемых боталовых протоков (см. рис. 77 на с. 137).
У хвостатых амфибий, у которых пожизненно сохраняются жабры, схема кровообращения очень близка к таковой у рыб и личинок высших амфибий. У них от брюшной аорты отходят четыре пары артериальных дуг. Сонные артерии отходят от выносящей части первой дуги и сохраняются сонные протоки . С появлением же легких образуются легочные артерии, берущие начало от четвертой артериальной дуги. Схема кровообращения в этом случае почти тождественна таковой двоякодышащих рыб (см. рис. 44 на с. 83).
Венозная система низших амфибий похожа на венозную систему двоякодышащих рыб. Хвостовая вена делится на две воротные вены почек. Из них кровь поступает в непарную заднюю полую вену и в парные задние кардинальные вены . Последние на уровне сердца сливаются с парными яремными, подключичными и кожными венами и образуют кювьеровы протоки, изливающие кровь в венозную пазуху. От кишечника кровь собирается по подкишечной и брюшной венам , которые, сливаясь, образуют воротную вену печени . Из печени кровь поступает в полую вену по печеночной вене.
У бесхвостых амфибий кардинальные вены не сохраняются, и вся кровь из туловищной области собирается в конечном счете в заднюю полую вену , впадающую в венозную пазуху. Как и в предыдущем случае, имеются брюшная и подкишечная вены, образующие воротную систему кровообращения в печени. В связи с отсутствием

Рис. 74. Схема строения вскрытого сердца лягушки:
1 - правое предсердие; 2 - левое предсердие; 3 - желудочек; 4 - клапан, закрывающий общее отверстие, ведущее из обоих предсердий в желудочек; 5 -артериальный конус; 6 - общий артериальный ствол; 7 - кожно-легочная артерия; 8 - дуга аорты; 9 - общая сонная артерия; 10 - сонная "железа"; 11 - спиральный клапан артериального конуса
кардинальных вен у бесхвостых не образуются и кювьеровы протоки. Яремные вены , сливаясь с подключичными, образуют в этом случае парные передние полые вены , впадающие в венозный синус , или пазуху. В верхние полые вены впадают и кожные вены соответствующей стороны, которые несут не венозную, а артериальную кровь.
Легочные вены несут кровь непосредственно в левое предсердие. Венозная кровь (с довольно значительной примесью окисленной крови, поступающей из кожных вен в передние полые вены) изливается в венозную пазуху (синус), а оттуда в правое предсердие. При сокращении предсердий венозная и артериальная кровь изливается через общее для обоих предсердий отверстие в желудочек. К его правой части примыкает артериальный конус, в который поступает сначала более венозная кровь, направляющаяся далее в открытое отверстие кожно-легочных артерий. Отверстия остальных артериальных дуг в это время прикрыты спиральным клапаном артериального конуса . При дальнейшем сокращении желудочка давление в артериальном конусе возрастает, спиральный клапан сдвигается и открываются отверстия системных дуг аорты, через которые поступает смешанная кровь из центральной части желудочка. Дальнейшее сдвигание спирального клапана освобождает устья сонных артерий, куда и проходит наиболее окисленная кровь, выходящая в артериальный конус в последнюю очередь из левой части желудочка. При всем этом полного разделения потоков артериальной и венозной крови все же нет.
Скорость движения крови (важный показатель интенсивности метаболизма) у амфибий невелика. Об этом косвенно свидетельствует частота пульса. У травяной лягушки с массой тела 50 г она равна 40-50 ударам в минуту. Для сравнения укажем, что у сходной по размерам птицы этот показатель равен примерно 500. У водных форм наблюдается брадикардия. Уровень артериального давления у амфибий также низок. У хвостатых он равен примерно 22/12-30/25, у бесхвостых - 30/20. Для сравнения: у чешуйчатых рептилий этот показатель составляет около 80/60 (Проссер, 1978).
Нервная система . Головной мозг (рис. 75) характеризуется рядом прогрессивных черт. Это выражается в относительно более крупных, чем у рыб, размерах переднего мозга, в полном разделении его полушарий и в том, что не только дно боковых желудочков, но и их бока и крыша содержат нервные клетки. Таким образом, у земноводных имеется настоящий мозговой свод - архипаллиум , который среди костных рыб свойствен только двоякодышащим. Средний мозг сравнительно небольших размеров. Мозжечок очень мал, а у некоторых хвостатых (у протеев) он практически

Рис. 75. Головной мозг лягушки:
I - сверху, II - снизу; III - сбоку; IV - в продольном разрезе; 1 - полушария переднего мозга; 2 - обонятельная доля, 3 - обонятельный нерв; 4 - промежуточный мозг; 5 - зрительная хиазма; 6 - воронка, 7 - гипофиз, 8 - средний мозг; 9 - мозжечок; 10 - продолговатый мозг; 11 - четвертый желудочек, 12 – спинной мозг 13 - третий желудочек; 14 - сильвиев водопровод; 15 - головные нервы
незаметен. Слабое развитие этой части головного мозга связано с крайне однообразными, несложными движениями амфибий. Завершает все продолговатый мозг . От головного мозга отходят десять пар головных нервов (1-Х).
Спинномозговые нервы у хвостатых и бесхвостых образуют хорошо выраженные плечевое и поясничное сплетения. Хорошо развита симпатическая нервная система , представленная в основном двумя нервными стволами, расположенными по бокам позвоночника.
Органы чувств у земноводных более развиты, чем у рыб. Органы обоняния амфибий представляют парные обонятельные капсулы , внутренняя поверхность которых выстлана обонятельным эпителием . Они сообщаются с наружной средой парными наружными ноздрями; от обонятельных капсул отходят внутренние ноздри (хоаны), сообщающиеся с ротоглоточной полостью. У амфибий, как и у всех наземных позвоночных, указанная система служит для восприятия запахов и для дыхания.
Органы боковой линии
Органы вкуса . Расположены в ротовой полости. Предполагается, что лягушка воспринимает только горькое и соленое.
Органы зрения . Глаза амфибий имеют ряд особенностей, связанных с полуназемным образом жизни: 1) подвижные веки защищают глаза от высыхания и загрязнения; при этом кроме верхнего и нижнего век имеется еще третье веко, или мигательная перепонка , расположенная в переднем углу глаза; 2) есть слезная железа , секрет которой омывает глазное яблоко; 3) выпуклая (а не плоская, как у рыб) роговица и линзовидный (а не круглый, как у рыб) хрусталик ; обе последние особенности определяют более дальнозоркое зрение амфибий (интересно, что в воде роговица у амфибий становится плоской); 4) аккомодация зрения достигается, как у акул, смещением хрусталика под действием ресничного мускула. О цветовом зрении амфибий информации нет.
Орган слуха устроен значительно более сложно, чем у рыб, и приспособлен к восприятию звуковых раздражений в воздушной среде. Наиболее полно это выражено у бесхвостых амфибий. Кроме внутреннего уха , представленного, как и у рыб, перепончатым лабиринтом , у земноводных имеется еще среднее ухо . Последнее представляет собой полость, один конец которой открывается в ротоглотку, а другой подходит к самой поверхности головы и затянут тонкой перепонкой, которая называется барабанной . Полость эта образует изгиб, вершина которого находится у перепончатого лабиринта. Верхняя часть полости от барабанной перепонки до перепончатого лабиринта носит название барабанной полости . В ней расположена палочковидная косточка - стремя , которая одним концом упирается в овальное окно внутреннего уха, другим - в барабанную перепонку. Нижняя часть полости среднего уха, открывающаяся в ротоглотку, именуется евстахиевой трубой .
Данные сравнительной анатомии и эмбриологии показывают, что полость среднего уха гомологична брызгальцу рыб, т.е. рудиментарной жаберной щели, лежащей между челюстной и подъязычной дугами, а слуховая косточка гомологична верхнему отделу подъязычной дуги - гиомандибуляре. На этом примере видно, что важное изменение органа может достигаться путем видоизменения и смены функций образований, имевшихся ранее у примитивных форм.
У безногих и хвостатых барабанная перепонка и барабанная полость отсутствуют, но слуховая косточка хорошо развита. Редукция среднего уха у этих групп, видимо, явление вторичное.
Органы боковой линии свойственны личинкам всех амфибий. Во взрослом состоянии они сохраняются только у водных форм хвостатых амфибий и немногих, тоже водных, бесхвостых. В отличие от рыб, чувствующие клетки этого органа расположены не в углубленном канале, а лежат поверхностно в коже.
Органы выделения (рис. 76) устроены по типу их организации у хрящевых рыб. В зародышевом состоянии органом выделения
служит пронефрос , у взрослых - мезонефрос с его типичным выводным путем - волъфовым каналом . Мочеточники открываются в клоаку. Сюда же у высших наземных амфибий открывается мочевой пузырь . После его наполнения моча через то же отверстие выводится в клоаку и затем изгоняется наружу.
Число нефронов у низших (хвостатых) амфибий около 500, у высших (бесхвостых) -около 2000. Такая заметная разница, видимо, определяется тем, что у хвостатых, которые теснее связаны с водоемами, имеет место и внепочечный (через кожу и жабры) путь выделения продуктов азотистого обмена. У хвостатых, в отличие от бесхвостых, нефроны (или хотя бы часть их) имеют нефростомы, т.е. воронки, сообщающие их с полостью тела (примитивный признак). Сосудистые клубки в боуменовых капсулах развиты хорошо, и амфибии выделяют много жидкой мочи. Укажем для примера, что у лягушек рода Rana скорость фильтрации крови составляет около 35 мл/(кг*ч).
Основным продуктом белкового обмена у амфибий служит мочевина, которая не очень токсична, но для выведения из организма требует большого количества воды, в которой она растворяется. Физиологически это вполне оправдано, так как поглощение воды организмом у амфибий в подавляющем случае не встречает затруднений.
Связь типа белкового обмена с условиями внешней среды доказывается и следующими двумя примерами. У тритона осенью на суше доля аммиака в общих продуктах азотистого обмена равна 13%, а при летнем водном существовании она возрастает до 26%. У головастика доля аммиака равна 75%, а у лягушонка, утерявшего хвост, с развитыми ногами - всего 16%.
Органы размножения . У самцов парные семенники не имеют самостоятельных выводных путей. Семявыносящие канальцы

Рис. 76. Мочеполовая система самца лягушки:
1 - почка; 2 - мочеточник; 3 - полость клоаки; 4 - мочеполовое отверстие; 5 - мочевой пузырь; 6 - отверстие мочевого пузыря; 7 - семенник; 8 - семявыно-сящие каналы, 9 - семенной пузырек; 10 - жировое тело, 11 - надпочечник
проходят через передний отдел почки и впадают в вольфов канал, который, таким образом, служит не только мочеточником, но и семявыносящим каналом. Каждый вольфов канал у самцов перед впадением в клоаку образует расширение - семенной пузырек , в котором временно резервируется семя.
Над семенниками лежат жировые тела - образования неправильной формы желтого цвета. Они служат для питания семенников и развивающихся в них сперматозоидов. Величина жировых тел меняется по сезонам. Осенью они велики; весной же, во время интенсивного сперматогенеза, вещество их энергично расходуется и размеры жировых тел резко сокращаются. Копулятивных органов у подавляющего большинства амфибий нет.
У самок развиваются парные яичники , над ними также лежат жировые тела. Созревшие яйца попадают в полость тела, откуда они поступают в воронкообразные расширения парных яйцеводов - мюллеровых каналов . Яйцеводы - длинные, сильно извитые трубки, задний отдел которых открывается в клоаку.
Из изложенного видно, что, как и у хрящевых рыб, у самцов земноводных мочевой и половой проток объединены и представляют собой единый вольфов канал, у самки же вольфов канал выполняет функцию только мочеточника, а половые продукты выводятся через самостоятельный половой проток - яйцевод, или мюллеров канал.
Земноводные, или амфибии, во взрослом состоянии, как правило, наземные животные, но они еще тесно связаны с водной средой, а их личинки постоянно обитают в воде. Следовательно, русское и греческое (амфибиос - ведущий двойную жизнь) названия отражают главную черту этих позвоночных. Земноводные произошли, как указывалось выше, от девонских кистеперых рыб, обитавших в небольших пресных водоемах и выползавших при помощи своих мясистых парных плавников на берег.
Наружное строение.
Тело (рис. 147) состоит из головы, туловища, передних и задних парных расчлененных конечностей. Конечности состоят из трех отделов: передние - из плеча, предплечья и кисти, задние - из бедра, голени и стопы. Хвост имеется лишь у меньшинства современных земноводных (отряд хвостатые - тритоны, саламандры и др.). Он редуцирован у взрослых форм самой большой группы земноводных - бесхвостых (лягушки, жабы и др.) в связи с приспособлением последних к передвижению прыжками по суше, но сохранился у их личинок - головастиков, обитающих в воде. У немногих видов, ведущих полуподземный образ жизни (отряд безногие, или червяги) редуцировались конечности и хвост.
Голова подвижно сочленяется с туловищем, хотя движение ее очень ограничено и выраженной шеи нет. Расчлененные конечности и подвижное соединение головы с туловищем - характерные признаки наземных позвоночных
, у рыб они отсутствуют. Тело наземных форм сплющено в дорзо-вентраль-ном направлении, в то время как у рыб (в связи с приспособлением их к плаванию) оно, как правило, сдавлено с боков. У водных амфибий форма тела приближается к рыбообразной. Величина тела колеблется от 2 до 160 см (японская саламандра); в среднем амфибии имеют меньшую величину, чем остальные наземные животные. Кожа голая, богатая железами, отделенная во многих местах от мускулатуры благодаря наличию подкожных лимфатических полостей. Она снабжена большим количеством кровеносных сосудов и выполняет также дыхательную функцию (см. далее). У некоторых видов выделения кожных желез ядовиты. Окраска кожи очень разнообразна.
Нервная система.
В связи с приспособлением амфибий к жизни на суше и в особенности в связи с коренным изменением характера передвижения нервная система довольно сильно изменилась. Передний мозг у земноводных (см. рис. 133,В) большей величины, чем средний; у рыб, как правило, наблюдается обратное соотношение. Это объясняется тем, что у рыб функции переднего мозга связаны только с восприятием обонятельных раздражений, у земноводных он начинает принимать участие в координации разных функций тела и в его поверхностном слое появляются зачатки коры (пока еще очень слабые), в которой сосредоточены нервные клетки. В то же время следует отметить, что в переднем мозге хорошо развиты обонятельные доли. Мозжечок у амфибий развит в отличие от рыб очень слабо. Рыбы постоянно движутся, причем положение их тела неустойчивое, земноводные же, опираясь на ноги, находятся в довольно устойчивом положении. Участки спинного мозга, в местах отхождения от него нервов, идущих к мышцам ног, которые выполняют значительно большую работу, чем мышцы парных плавников рыб, утолщены и с ними связаны плечевое и поясничное сплетения нервов. Периферическая нервная система сильно изменилась в связи с дифференциацией мускулатуры (см. дальше) и появлением длинных, членистых конечностей.

Из органов чувств наиболее существенным изменениям подвергся орган слуха. Передача звуковых волн из водной среды в ткани животного, которые тоже насыщены водой и имеют приблизительно такие же акустические свойства, как и вода, происходит значительно лучше, чем из воздуха. Звуковые волны, распространяющиеся в воздушной среде, почти Целиком отражаются от поверхности животного и только около 1 % энергии этих волн проникает в его тело. В связи с этим у земноводных возник в добавление к лабиринту, или внутреннему уху, новый отдел органа слуха - среднее ухо. Оно представляет собой (рис. 148) наполненную воздухом маленькую полость, сообщающуюся с ротовой полостью посредством евстахиевой трубы и закрытую снаружи тонкой, Упругой барабанной перепонкой. В среднем ухе находится слуховая досточка (или столбик), которая одним концом упирается в барабанную перепонку, а другим - в окошечко, затянутое пленкой и ведущее в полость черепа, где находится лабиринт, окруженный перилимфой. Давление внутри среднего уха равно атмосферному и барабанная перепонка может вибрировать под влиянием звуковых воздушных волн, воздействие которых дальше через слуховую косточку и перилимф передается стенкам лабиринта и воспринимается окончаниями слухового нерва. Полость среднего уха образовалась из первой жаберной щели, а столбик - из расположенной около щели подъязычнонижнечелюстной кости (гиомандибуляре), которая подвешивала висцеральный отдел черепа к мозговому там, где за ушными костями находился лабиринт.

Глаза прикрыты подвижными веками, которые защищают органы зрения от высыхания и засорения. Благодаря изменению формы роговицы и хрусталика земноводные видят дальше, чем рыбы. Амфибии хорошо воспринимают небольшие температурные изменения. Они чувствительны к воздействиям различных веществ, растворенных в воде. Их орган обоняния реагирует на раздражения, вызываемые газообразными веществами. Таким образом, органы чувств земноводных претерпели ряд изменений в связи с переходом к обитанию на суше. У личинок и взрослых животных, обитающих постоянно в воде, имеются, как и у рыб, органы боковой линии.
Земноводным свойственны довольно сложные инстинктивные действия, особенно в период размножения. Например, самец жабы-повитухи, обитающей в России на западе Украины, наматывает на свои задние конечности «шнуры» икры и до развития головастиков прячется в укромных местах на берегу. Через 17-18 дней самец возвращается в воду, где происходит вылупление головастиков. Это своеобразный инстинкт защиты потомства. У ряда тропических бесхвостых амфибий известны еще более сложные инстинкты. Земноводным свойственны и условные рефлексы, однако они вырабатываются с большим трудом.
Двигательная система и скелет. Мышечная система в связи с различными приспособлениями к жизни на суше (развитие конечностей наземного типа, возникновение подвижного сочленения головы с туловищем и др.) подверглась коренным преобразованиям, хотя и сохранила много особенностей, присущих рыбам. Мышечная система рыб очень однообразна и в основном состоит из сходных боковых мышечных сегментов. У земноводных мышечная система стала более дифференцированной, состоящей из разнообразных мышц (рис. 149). У амфибий заложены основы той мышечной системы, которая потом развивалась и усложнялась у настоящих сухопутных позвоночных - пресмыкающихся, птиц и млекопитающих. Это относится и к скелету.
Череп амфибий имеет много хрящевых элементов, что, вероятно, объясняется необходимостью облегчить в связи с полуназемным образом жизни массу тела. В составе черепа много костей, перечисленных при описании черепа высших рыб, и в том числе характерный только для рыб и земноводных парасфеноид (рис. 150). Так как гиомандибуляре стала слуховой косточкой, то роль подвеска выполняет квадратная кость. В связи с утратой во взрослом состоянии жаберного аппарата жаберные дуги редуцируются и сохраняются лишь их видоизмененные остатки. Подъязычная дуга сильно изменяется и частично редуцируется. Череп амфибий очень широкий, что отчасти связано с особенностями их дыхания. Нижняя челюсть, как и у костных рыб, состоит из нескольких костей.
Позвоночный столб (рис. 150) у бесхвостых очень короткий и заканчивается длинной косточкой - уростилем, образовавшимся из рудиментов хвостовых позвонков. У хвостатых земноводных хвостовой отдел позвоночного столба состоит из ряда позвонков. У этих амфибий хвост играет существенную роль при передвижении: в воде он служит для плавания, на суше - для поддержания равновесия. Ребра слабо развиты (у хвостатых земноводных) или редуцированы, а их остатки слились с поперечными отростками позвонков (у остальных амфибий), у древних амфибий ребра имелись. Редукция их у современных форм объясняется необходимостью облегчить массу тела (сильно возросшую при переходе из водной среды в воздушную) у этих позвоночных, еще недостаточно приспособленных к передвижению по суше. Вследствие редукции ребер у земноводных нет грудной клетки. Первый позвонок устроен иначе, чем у рыб: он имеет две суставные впадины для сочленения с двумя затылочными мыщелками черепа, благодаря чему голова земноводных стала подвижной.
Скелет передней конечности (рис. 150) состоит из плечевой кости, двух костей предплечья - лучевой и локтевой, костей запястья, пястных костей и фаланг пальцев. Скелет задней конечности (рис. 150) состоит из бедра, двух костей голени - большой берцовой и малой берцовой, костей предплюсны, плюсневых костей и фаланг пальцев. Следовательно, сходство в строении обеих пар конечностей, несмотря на некоторые различия в их функциях, очень велико. Первоначально передние и задние ноги были пятипалые, у современных земноводных число пальцев может быть меньшим. Задние конечности у многих бесхвостых амфибий служат и для плавания, в связи с чем они удлинились, а пальцы соединены плавательными перепонками.
Пояса конечностей развиты значительно лучше, чем у рыб. Плечевой пояс состоит из костных и хрящевых элементов: лопатки, ключицы, вороньей кости (коракоида) и др. (рис. 150). Ключицы и коракоиды соединены с грудиной, в состав которой тоже входят костные и хрящевые элементы. Головка плеча сочленяется с плечевым поясом. Задний пояс конечностей, или таз, состоит из трех костей: подвздошной, лобковой и седалищной (рис. 150). Образуемая этими костями большая вертлужная впадина служит для сочленения с головкой бедра. Таз соединен с одним позвонком - крестцовым, благодаря чему задние ноги в отличие от брюшных плавников рыб получили довольно прочную опору.
Кровеносная система. У личинок земноводных, обитающих в воде и дышащих жабрами, кровеносная система в основных чертах сходна с кровеносной системой рыб, у взрослых же животных, ведущих наземный образ жизни, она существенно меняется в связи с заменой жаберного дыхания легочным, усилением кожного дыхания, развитием конечностей сухопутного типа и другими изменениями тела. Сердце (см. рис. 134, В, 151) состоит из трех камер: правого и левого предсердий и одного желудочка. От правой части последнего отходит артериальный конус (он был и у рыб, предков земноводных), от которого берут начало четыре пары артерий: первая пара - сонные артерии , несущие кровь в голову, вторая и третья пары - сосуды, соединяющиеся в самый большой сосуд тела - аорту , разветвления которой направляются в разные части тела, четвертая пара - легочнокожные артерии , которые потом разделяются на самостоятельные кожные и легочные артерии.
Из легких кровь, насыщенная кислородом, через легочные вены попадает в левое предсердие, а кровь, насыщенная во всех частях тела углекислым газом, в передней части тела вливается в переднюю полую вену, а в задней части тела - в заднюю полую вену (рис. 152). Обе полые вены впадают в венозный синус , откуда кровь (насыщенная углекислым газом) вливается в правое предсердие. Из обоих предсердий кровь поступает в единственный желудочек сердца. Внутренняя поверхность желудочка имеет углубления и поэтому кровь в нем не успевает полностью смешаться: в левой части его - кровь, насыщенная кислородом, в правой части - кровь, насыщенная углекислым газом, а в средней части - смешанная. Поскольку артериальный конус начинается с правой стороны желудочка, то первая порция крови, попадающая в него (т. е. артериальный конус), будет венозная, она направляется в самые задние артерии - легочные.
Идущая затем смешанная кровь попадает в артерии, образующие аорту, а через разветвления последней во все части тела. Насыщенная кислородом кровь из левой части желудочка направляется в сонные артерии. К этому нужно добавить, что кровь, насыщенная в коже кислородом, попадает, как уже отмечалось выше, через переднюю полую вену и венозный синус в правое предсердие и таким образом разбавляет находящуюся там венозную кровь, далее выталкиваемую в сосуды, образующие аорту. Следовательно, благодаря описанным выше приспособлениям, а также другим, не описанным здесь, разные части тела получают кровь, неодинаково насыщенную кислородом. На рис. 152 изображены главные артериальные и венозные сосуды амфибий.
У земноводных в связи с сильным развитием конечностей и большим, чем у рыб, расчленением тела, существенно изменилась сеть кровеносных сосудов. Появилось много новых сосудов, отсутствовавших у рыб, и сложилась система сосудов, характерная для наземных позвоночных. В то же время следует помнить, что кровеносная система амфибий значительно проще, чем у высших позвоночных.
Дыхательная система. Почти у всех земноводных есть легкие (см. рис. 151; 153). Эти органы имеют еще очень простое строение и представляют собой тонкостенные мешки, в стенках которых разветвляется довольно густая сеть кровеносных сосудов. Так как внутренняя стенка легких почти гладкая, то их поверхность относительно невелика. Трахея почти неразвита и легкие связаны непосредственно с гортанью. Поскольку грудной клетки у земноводных нет (см. выше), акт дыхания обеспечивается работой мышц ротовой полости. Вдох происходит следующим образом. При открытых ноздрях (которые в отличие от ноздрей рыб сквозные, т. е. кроме наружных ноздрей имеются и внутренние ноздри - хоаны ) и закрытом рте оттягивается дно обширной ротовой полости и в нее поступает воздух. Затем ноздри закрываются особыми клапанами, дно ротовой полости поднимается и воздух нагнетается в легкие. Выдох происходит в результате сокращения брюшной мускулатуры.
Значительное количество кислорода земноводные получают через кожу и слизистые оболочки ротовой полости. У некоторых видов саламандр легких совсем нет и весь газообмен совершается через кожу. Однако кожа только в том случае может выполнять функции дыхания, если она влажная. Поэтому обитание земноводных в условиях недостаточной для них влажности воздуха невозможно. Личинки, живущие в воде, дышат жабрами (сначала - наружными, потом - внутренними) и кожей. У некоторых хвостатых земноводных, постоянно живущих в воде, жабры сохраняются всю жизнь. Таким образом, по способам дыхания амфибии еще близки к рыбам.
Выделительная система. Почки (см. рис. 136,А, Б; рис. 151), как и у рыб, туловищные. Вольфовы каналы впадают в клоаку. Туда же открывается мочевой пузырь, в котором накапливается моча. Удаление продуктов диссимиляции происходит, кроме того, через кожу и легкие.
Пищеварительная система. Ротовая полость очень широкая. У ряда видов (главным образом у хвостатых амфибий) имеется много мелких, однородных, примитивно устроенных зубов, которые сидят на челюстях, сошнике, небных и других костях и служат лишь для удержания добычи. У большинства видов (главным образом у бесхвостых амфибий) зубы частично или полностью редуцируются, зато у них сильно развивается язык. Последний у лягушек прикреплен передним концом и может для поимки добычи далеко выбрасываться задним концом вперед. Он очень липкий и хорошо приспособлен для выполнения указанной функции. У видов, постоянно живущих в воде, язык обычно редуцируется. Захват добычи у таких земноводных осуществляется челюстями.
Пищеварительная трубка (см. рис. 151) сравнительно короткая и состоит из глотки, пищевода, желудка, тонкой кишки и совсем небольшой прямой (толстой) кишки. Задняя часть прямой кишки представляет собой клоаку; через нее выводятся кроме кала моча и половые продукты. В ротовую полость впадают слюнные железы, отсутствующие у рыб. Секрет этих желез служит главным образом для смачивания пищи. Слюнные железы очень слабо развиты у видов, обитающих в воде, и значительно лучше - у наземных. Печень велика; поджелудочная железа хорошо выражена. Пища взрослых земноводных в основном животная (насекомые, мелкие позвоночные и др.). Головастики же бесхвостых амфибий большей частью растительноядны.

Размножение. Мужские половые железы (семенники) лежат около почек (см. рис. 151, Б). Их протоки открываются в канальцы передней части почек (см. рис. 136, А) и семя выводится, как и моча, через вольфовы каналы. Женские половые железы (яичники) сильно разрастаются в период икрометания. Икра выходит через очень длинные мюллеровы каналы (см. рис. 136, Б). Последние не имеют прямой связи с яичниками и созревающая икра попадает через полость тела в воронки мюллеровых каналов.
Оплодотворение в большинстве случаев происходит в воде. У многих амфибий этому предшествует сближение самца и самки: самец обхватывает сзади самку, надавливает передними конечностями на ее брюшную стенку и этим способствует выходу в воду икры, которую он сразу оплодотворяет. Таким образом, при наличии полового акта оплодотворение происходит вне организма самки. У меньшинства видов (например, у тритонов) самец выделяет семя в особом мешочке (сперматофоре), который самка тут же захватывает краями клоаки. В данном случае полового акта нет, но оплодотворение - внутреннее. Наконец, у некоторых видов самец вводит семя в клоаку самки при помощи своей способной выпячиваться клоаки.
У многих видов хорошо выражен половой диморфизм (в окраске, строении передних ног, которыми самцы удерживают самок, и по другим признакам). Самцы ряда видов могут издавать очень громкие звуки благодаря усилению этих звуков голосовыми мешками - резонаторами.
Развитие. Развитие земноводных, как правило, происходит в воде. Из оплодотворенных яиц развиваются личинки (головастики), имеющие рыбообразную форму. Дышат они жабрами и их внутреннее строение напоминает рыб. В течение периода роста происходит превращение (метаморфоз) головастиков: сначала у них вырастают задние ноги, потом передние, жабры и хвост атрофируются (у бесхвостых), развиваются легкие, большие изменения происходят в кровеносной системе и т. д.
Происхождение. Земноводные, как было разъяснено выше (с. 296), произошли от кистеперых рыб. Парные плавники древних кистеперых рыб, из которых развились членистые конечности наземных позвоночных, были коротки и широки, в их состав входило множество мелких костных элементов, не соединенных суставами, расположенных в несколько (не менее восьми) поперечных рядов. Пояса, на которые опирались плавники, были сравнительно слабо развиты (особенно тазовый пояс). В связи с превращением плавников в конечности наземного типа произошли существенные изменения в скелете.
Во-первых, редуцировались многие костные элементы: в первых проксимальных рядах осталось по одной кости, в передней ноге - плечо, в задней - бедро; во вторых рядах - по две кости, в передней ноге - лучевая и локтевая кости, в задней - большая и малая берцовые кости; в двух последующих рядах осталось по пять костей, в передней ноге образовавших запястье, в задней - предплюсну; в следующем ряду оставшиеся пять костей вошли в передней ноге в состав пястья, в задней - плюсны; остальные три ряда с пятью костями в каждом стали фалангами пальцев. Уменьшение количества костей способствовало усилению прочности ног.
Во-вторых, кости двух первых рядов (т. е. плеча и предплечья, бедра и голени) сильно удлинились, что было очень важно для усиления быстроты движения.
В-третьих, между перечисленными костями развились суставы, т. е. конечности стали членистыми, что является важнейшим условием их работы.
В-четвертых, укрепились пояса конечностей (см. выше описание поясов). Параллельно указанным изменениям произошли глубокие изменения в нервной, мышечной и сосудистой системах ног. Об изменении других систем органов, происшедших при превращении кистеперых рыб в амфибий, рассказано в общей характеристике последних.
Самыми древними земноводными были стегоцефалы (рис. 154), которые были многочисленны в каменноугольном периоде и окончательно вымерли в триасе. Они жили по берегам водоемов и проводили много времени в воде. Голова этих животных была покрыта щитками, чем и объясняется их название (стегоцефалы - покрытоголовые). В их скелете было много примитивных черт: костные элементы ног были малы и слабо отличались между собой по величине, позвонки двояковогнутые, пояса конечностей плохо развиты и т. д. От них произошли современные группы амфибий.